すべての染色体障害のうち、染色体数の異常は、核造影から最も明らかに識別可能である。 染色体数の障害には、染色体全体の複製または喪失、ならびに染色体の完全なセットの数の変化が含まれる。 それらは、相同染色体または姉妹染色分体の対が減数分裂の間に分離できないときに起こる非分裂によって引き起こされる。 ずれや不完全なシナプス、または染色体の移動を容易にする紡錘装置の機能不全は、非接合を引き起こす可能性があります。 非解離のリスクは、両親の年齢とともに増加する。
減数分裂IまたはIIのいずれかの間に非分裂が起こる可能性があり、結果は異なります(図1)。 相同染色体が減数分裂Iの間に分離できない場合、その結果、その特定の染色体を欠く2つの配偶子と、2つのコピーの染色体を持つ2つの配偶子が 減数分裂IIの間に姉妹染色分体が分離できない場合、その結果、その染色体を欠く1つの配偶子、1つの染色体のコピーを持つ2つの正常な配偶子、およ
練習問題
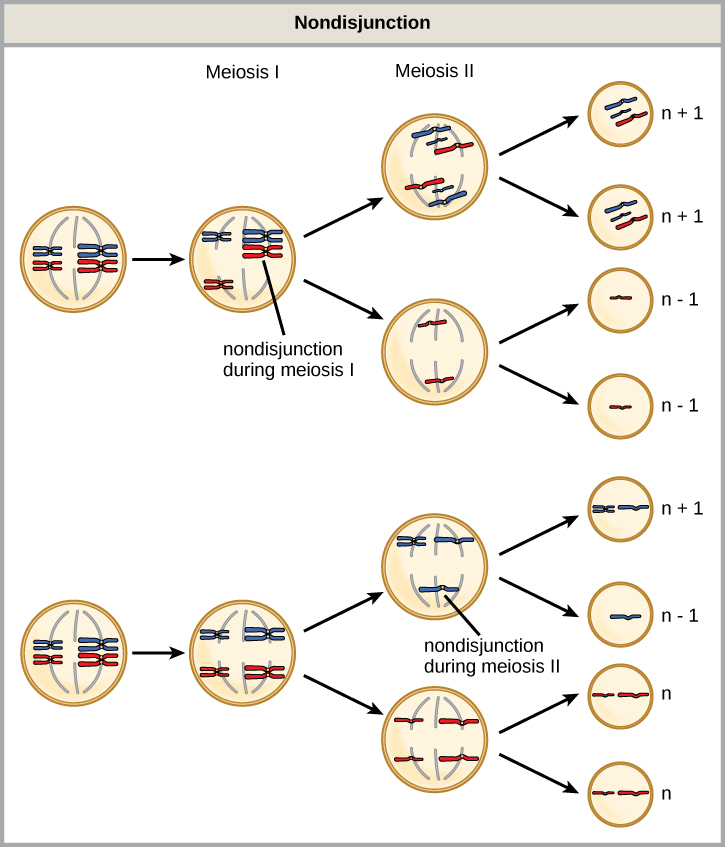
図1. 非分裂は、減数分裂中に相同染色体または姉妹染色分体が分離できず、染色体数が異常になると発生します。 減数分裂Iまたは減数分裂IIの間に非分裂が起こることがある。
非分離についての次の声明のどれが本当ですか?
- 非解離は、n+1またはn–1染色体を持つ配偶子のみをもたらします。
- 減数分裂IIの間に発生する非分裂は、50パーセントの正常な配偶子をもたらす。
- 減数分裂中の非分裂Iは50パーセントの正常な配偶子をもたらす。
- 非解離は常に4つの異なる種類の配偶子をもたらします。
異数性

図2。 トリソミー21を有する胎児を有する発生率は、母体の年齢とともに劇的に増加する。
その種に適した数の染色体を持つ個体は正倍数体と呼ばれ、ヒトでは正倍数性は22対の常染色体と1対の性染色体に相当する。 染色体数の誤差を有する個体は、異数体、モノソミー(1つの染色体の喪失)またはトリソミー(外来染色体の獲得)を含む用語として記述される。 常染色体のいずれかのコピーを欠いている単系統ヒト接合体は、必須遺伝子を欠いているため、常に出生に発展することができません。 これは、ヒトにおける「遺伝子投与量」の重要性を強調している。 ほとんどの常染色体トリソミーはまた、出生に発展するために失敗します;しかしながら,小さな染色体のいくつかの重複(13,15,18,21,または22)数週間から多 トリソミック個体は、異なるタイプの遺伝的不均衡に苦しんでいる:遺伝子用量の過剰。 余分な染色体を持つ個体は、その染色体によってコードされる遺伝子産物の豊富さを合成することができる。 特定の遺伝子のこの余分な用量(150%)は、多くの機能的課題をもたらし、しばしば発達を妨げる可能性がある。 生存可能な出生の中で最も一般的なトリソミーは、ダウン症候群に対応する染色体21のトリソミーである。 この遺伝性障害を有する個体は、低身長および発育不全の数字、広範な頭蓋骨および大きな舌を含む顔面の区別、および有意な発達遅延を特徴とする。 ダウン症候群の発生率は母親の年齢と相関しており、高齢の女性はtrisomy21遺伝子型を持つ胎児を妊娠する可能性が高くなります(図2)。
倍数性

図3を示しています。 多くの倍数体植物と同様に、この三倍体オレンジdaylily(Hemerocallis fulva)は特に大きく、堅牢であり、その二倍体の対応の花びらの三倍の数で花を育てます。 (クレジット:スティーブ-カーグ)
染色体セットの正しい数(二倍体種のための二つ)よりも多くを持つ個体は倍数体と呼ばれています。 例えば、異常な二倍体の卵と正常な一倍体の精子との受精は、三倍体の接合体をもたらすであろう。 倍数体動物は、扁形動物、甲殻類、両生類、魚、トカゲの中で唯一のいくつかの例で、非常にまれです。 倍数体動物は減数分裂が正常に進行することができず、代わりに生存可能な接合体を得ることができない主に異数体の娘細胞を産生するため、滅菌 まれに、倍数体動物は、未受精卵が有糸分裂して子孫を産生する一倍体によって無性生殖することができる。 対照的に、倍数性は植物界では非常に一般的であり、倍数体植物はその種の正倍数体よりも大きく、より堅牢である傾向がある(図3)。
ヒトにおける性染色体非解離
ヒトは常染色体トリソミーおよびモノソミーで劇的な有害な影響を示す。 したがって、x染色体の数が異なるにもかかわらず、人間の女性と男性が正常に機能できることは直感に反するように見えるかもしれません。 常染色体の利得または喪失ではなく、性染色体の数の変動は比較的軽度の影響と関連している。 部分的には、これはX不活性化と呼ばれる分子プロセスのために起こる。 発達の初期に、雌の哺乳類胚がわずか数千の細胞(新生児の兆に比べて)で構成されている場合、各細胞の一つのX染色体は、バー体と呼ばれる静止(休眠)構造 X染色体(母系または父系)が各細胞で不活性化される可能性はランダムですが、不活性化が起こると、その細胞から誘導されたすべての細胞は同じ不活性X染色体またはBarr体を持つことになります。 このプロセスによって、女性はX染色体の彼らの二重遺伝の線量を償います。

図4。 猫では、コートの色の遺伝子はX染色体上に位置しています。 雌猫の胚発生では、2つのX染色体のうちの1つが各細胞で無作為に不活性化され、猫がコートの色に対して2つの異なる対立遺伝子を有する場合、べっ甲パターンが生じる。 オスの猫は、X染色体が一つしかないため、べっ甲のコートの色を示すことはありません。 (クレジット:マイケル-ボデガ)
いわゆる”べっ甲”猫では、胚のX不活性化は色の多様性として観察されます(図4)。 X-linked coat color遺伝子のヘテロ接合体である女性は、その領域の胚細胞前駆体で不活性化されたX染色体に対応して、体の異なる領域にわたって2つの異
異常な数のX染色体を持つ個体は、それぞれの細胞の1つのX染色体を除く全てを不活性化します。 しかし、不活性化されたX染色体でさえ、いくつかの遺伝子を発現し続け、X染色体は女性の卵巣の適切な成熟のために再活性化しなければならない。 その結果、X染色体異常は、典型的には、軽度の精神的および身体的欠陥、ならびに無菌性と関連している。 X染色体が完全に存在しない場合、個体は子宮内で発達しない。
性染色体数のいくつかの誤りが特徴づけられている。 Triplo-Xと呼ばれる三つのX染色体を持つ個人は、表現型の女性ですが、発達の遅れと生殖能力の低下を表現します。 一つのタイプのクリネフェルター症候群に対応するXXY遺伝子型は,小さな精巣,拡大した胸,および減少した体毛を有する表現型的に男性個体に対応する。 より複雑なタイプのKlinefelter症候群が存在し、その個体は5つのX染色体を有する。 すべてのタイプにおいて、1つを除くすべてのX染色体は、過剰な遺伝的投与量を補うために不活性化を受ける。 これは、各細胞核内のいくつかのBarr体として見ることができます。 ターナー症候群は、X0遺伝子型として特徴付けられる(すなわち、、単一の性染色体だけ)、低い身長、首の地域の水かきをされた皮、ヒアリングおよび心臓減損および生殖不能症の表現型の女性の個人に対応します。
重複および欠失
染色体全体の損失または利得に加えて、染色体セグメントが重複または失われる可能性があります。 重複や欠失は、多くの場合、生き残るが、物理的および精神的な異常を示す子孫を生成します。 複製された染色体セグメントは、既存の染色体に融合してもよく、または核内で遊離してもよい。 Cri-du-chat(フランス語で”猫の叫び”から)は、5p(5番染色体の小さな腕)の大部分の欠失に起因する神経系の異常および識別可能な身体的特徴に関連するシ この遺伝子型を有する乳児は、障害の名前が基づいている特徴的な甲高い叫び声を発する。

図5。 Cri-du-chat症候群のこの個体は、2歳、4歳、9歳、および12歳で示されています。 (クレジット:Paola Cerruti Mainardi)