- Introduction
- Expérience 1
- Méthodes
- Participants
- Appareil et tâche
- Phases expérimentales et procédure
- Analyse des données
- Résultats
- Temps total de mouvement
- Synchronisation relative
- Discussion
- Expérience 2
- Méthodes
- Participants
- Appareil, Tâche, Phases expérimentales, Procédure et Analyse des données
- Résultats
- Temps total de mouvement
- Synchronisation relative
- Discussion
- Discussion générale
- Contributions des auteurs
- Financement
- Déclaration de conflit d’intérêts
- Notes de bas de page
Introduction
Vous êtes un golfeur passionné et vous voulez apprendre un nouveau coup. Comment procéderiez-vous ? Il y a de fortes chances que vous observiez quelqu’un (en direct, en vidéo, sur Youtube, etc.) qui sait comment effectuer ce tir, et vous essaierez de comprendre quoi faire et comment le faire. La recherche indique clairement que cette stratégie d’apprentissage est couronnée de succès parce qu’il a été démontré que l’observation favorise l’apprentissage d’une grande variété de compétences motrices (voir McCullagh et al., 1989; Hodges et coll., 2007; Vogt et Thomaschke, 2007; Ste-Marie et coll., 2012; Lago-Rodríguez et coll., 2014, pour des revues sur l’apprentissage observationnel). En effet, l’observation a beaucoup de points communs avec la pratique physique, qui est le premier déterminant de l’apprentissage de la motricité. Plus précisément, il a été démontré que des variables, telles que la quantité de pratique (Carroll et Bandura, 1990; Blandin, 1994), la fréquence de la connaissance des résultats (, Badets et Blandin, 2004, 2005; Badets et al., 2006), et le calendrier de pratique (Blandin et al., 1994; Wright et coll., 1997), affectent l’apprentissage par la pratique observationnelle et la pratique physique de manière similaire. Ces données ont conduit à la proposition que l’observation et la pratique physique utilisent des processus très similaires. Cette proposition est étayée par les résultats d’études de neuroimagerie qui ont montré qu’un ensemble de structures neuronales (y compris le cortex prémoteur, le lobule pariétal inférieur, le sillon temporal supérieur, la zone motrice supplémentaire, le gyrus cingulaire et le cervelet), également appelé « réseau d’observation d’action » (AON) (Kilner et al., 2009; Oosterhof et coll., 2010), est activé à la fois lorsque des individus effectuent une tâche motrice donnée et lorsqu’ils observent d’autres effectuer cette même tâche motrice (Grafton et al., 1997; Buccino et coll., 2001; Gallese et coll., 2002; Cisek et Kalaska, 2004; Frey et Gerry, 2006; Cross et coll. Il s’agit de l’un des plus grands groupes de l’histoire de l’art et de l’art., 2014).
L’observation favorise l’apprentissage de la motricité, mais qui devriez-vous observer pour apprendre ce nouveau coup de golf? Un expert qui maîtrise le tir vous aidera probablement à développer une référence sur ce qu’il faut faire et comment le faire, mais devriez-vous observer quelqu’un comme vous qui apprend ce tir et qui vous donne probablement une meilleure chance de détecter et d’apprendre des erreurs ou des changements de stratégie? La recherche a montré que l’observation à la fois d’un modèle qualifié (Martens et al., 1976; McCullagh et coll., 1989; Lee et coll., 1994; Al-Abood et al., 2001; Heyes et Foster, 2002; Hodges et coll., 2003; Bird et Heyes, 2005) et un modèle novice mène à un apprentissage significatif (Lee et White, 1990; McCullagh et Caird, 1990; Pollock et Lee, 1992; McCullagh et Meyer, 1997; Black et Wright, 2000; Buchanan et al., 2008; Buchanan et Dean, 2010; Hayes et coll., 2010). Cependant, des résultats récents de notre laboratoire ont montré que l’apprentissage observationnel d’une nouvelle motricité est amélioré après l’observation de modèles novices et experts plutôt que d’un modèle novice ou expert seul (Rohbanfard et Proteau, 2011; Andrieux et Proteau, 2013, 2014). Nous pensons que ce format d’observation » variable » conduit non seulement au développement d’une bonne représentation du mouvement (observation experte) mais également au développement de processus efficaces de détection et de correction des erreurs (observation novice).
Dans la présente étude, la question d’intérêt est simple mais importante. Lors de l’utilisation d’un calendrier d’observation variable, l’apprentissage sera-t-il meilleur lorsque les observateurs seront informés au préalable de la « qualité » de la performance qu’ils s’apprêtent à voir ou sera-t-il meilleur lorsque les observateurs seront laissés évaluer les performances avant de recevoir des commentaires? Informer les observateurs de ce qu’ils sont sur le point de voir peut leur permettre de choisir s’ils observeront pour imiter ou plutôt observer pour détecter une erreur, ou des faiblesses dans les performances du modèle, ce qui pourrait faciliter le développement de ces processus. Alternativement, le fait que les participants évaluent la qualité des performances qu’ils ont observées peut activer des processus cognitifs plus élaborés que lorsque ces informations sont transmises (par exemple, la détection et la reconnaissance d’erreurs ou l’évaluation d’une stratégie alternative), ce qui entraîne un meilleur apprentissage de la tâche.
La tâche que nous avons choisie a obligé les participants à modifier le schéma de synchronisation relatif qui a naturellement émergé des contraintes de la tâche (Collier et Wright, 1995; Blandin et al., 1999) à un nouveau modèle imposé de calendrier relatif. Cela ressemble à un changement de tempo lors de l’exécution d’un service au tennis ou d’un entraînement au golf (Rohbanfard et Proteau, 2011). Les participants ont observé deux modèles démontrant une grande variété de performances. Dans un groupe, les observateurs ont été informés avant chaque essai du niveau de qualité (performance experte, avancée, intermédiaire, novice ou débutante) de ce qu’ils s’apprêtaient à voir, tandis qu’un deuxième groupe d’observateurs n’a reçu les mêmes informations qu’après la fin de chaque essai d’observation.
Expérience 1
Méthodes
Participants
Quatre-vingt-dix étudiants droitiers (45 hommes et 45 femmes; âge moyen = 20,5 ans; SD = 0,9 ans) du Département de kinésiologie de l’Université de Montréal ont participé à cette expérience. Les participants étaient naïfs à l’égard du but de l’étude et n’avaient aucune expérience préalable de la tâche, et tous les participants se sont déclarés droitiers. Aucun des participants n’a signalé de troubles neurologiques et tous avaient une vision normale ou corrigée à la normale. Les participants ont rempli et signé des formulaires de consentement individuel avant de participer. Le Comité d’éthique de la recherche en sciences de la santé de l’Université de Montréal a approuvé cette expérience.
Appareil et tâche
L’appareil était similaire à celui utilisé par Rohbanfard et Proteau (2011). Comme illustré à la figure 1, il se composait d’une base en bois (45 × 54 cm), de trois barrières en bois (11 × 8 cm) et d’un bouton de démarrage encastré dans une cible (11 × 8 cm). La distance entre le bouton de démarrage et la première barrière était de 15 cm. Les distances des trois segments restants de la tâche étaient respectivement de 32, 18 et 29 cm. Les barrières étaient placées perpendiculairement à la base en bois au début de chaque essai, ce qui donnait un circuit micro-interrupteur fermé. Tous les microrupteurs étaient connectés à un ordinateur via le port d’E / S d’un convertisseur analogique–numérique (National Instruments, Austin, Texas, États-Unis), et un minuteur milliseconde était utilisé pour enregistrer à la fois le temps total de mouvement (TMT) et le temps nécessaire pour terminer chaque segment de la tâche (temps intermédiaires, ITs).
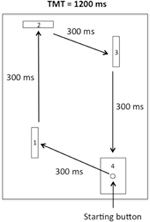
Figure 1. Croquis de l’appareil. Les participants ont dû quitter le bouton de démarrage et frapper les première, deuxième et troisième barrières dans le sens des aiguilles d’une montre avant d’atteindre finalement la cible.
Pour les essais de pratique physique (voir ci-dessous), les participants se sont assis près de la position de départ devant l’appareil. Ensuite, à partir du bouton de démarrage, les participants ont été invités à abattre successivement les première, deuxième et troisième barrières (libérant ainsi les microrupteurs) et à toucher finalement la cible dans le sens des aiguilles d’une montre comme illustré à la figure 1. Chaque segment de la tâche devait être complété dans un IT de 300 ms, pour un TMT de 1200 ms. Le schéma de mouvement, SON et TMT ont été illustrés sur une affiche située directement devant l’appareil pendant toutes les phases expérimentales.
Phases expérimentales et procédure
Les participants ont été assignés au hasard à l’un des trois groupes, chacun composé de 30 participants (15 femelles par groupe): contrôle (C), KR et observation (FW) et KR d’observation et de rétroaction (FB). Tous les groupes ont effectué quatre phases expérimentales, réparties sur 2 jours successifs.
Tous les participants ont reçu des instructions verbales concernant la TMT et les ITS avant la première phase expérimentale. La première phase expérimentale a été un pré-test, dans lequel tous les participants ont effectué 20 essais de pratique physique sans connaissance des résultats (KR) sur le TMT et le ITs.
La deuxième phase était une phase d’acquisition et consistait en 60 essais d’observation pour les participants des deux groupes d’observation (FW et FB). Ces participants ont regardé individuellement une présentation vidéo de deux modèles effectuant physiquement la tâche expérimentale. Pour chaque essai d’observation, KR concernant les performances du modèle (TMT et ITs) a été présenté en ms (voir Figure 1) soit avant la démonstration pour le groupe FW, soit après la démonstration pour le groupe FB. Le modèle a été modifié tous les cinq essais (c.-à-d. modèle 1 : essais 1 à 5 et modèle 2: essais 6 à 10, etc.), pour un total de 30 essais effectués par un modèle et de 30 essais effectués par l’autre modèle. Pour les groupes FW et FB, les deux modèles, qui ont participé à des travaux antérieurs de notre laboratoire, ont été choisis car pour les deux modèles, nous avions six clips vidéo illustrant les performances dans chacune des cinq sous-catégories. Ainsi, les participants des groupes FW et FB ne pouvaient associer un modèle particulier à une performance meilleure ou moins bonne. Une performance d’expert correspond à une erreur quadratique moyenne racine (RMSE ; voir la section analyse des données pour plus de détails sur le calcul) comprise entre 0 et 15 ms; les performances avancées, intermédiaires, novices et débutantes correspondaient à des RMSE de 30-45 ms, 60-75 ms, 90-105 ms et 120+ ms, respectivement. Les participants des groupes FW et FB ont été informés de la performance du modèle dans les États membres; ils ont également été informés du niveau de performance auquel il faisait référence. Les 30 essais résultants de chaque modèle (cinq niveaux de performance × six répétitions) ont été randomisés de sorte que les cinq niveaux de performance ont été présentés une fois dans chaque série de cinq essais. Pour éviter l’imitation physique de la séquence, qui pourrait interférer avec les processus d’observation, nous avons demandé aux participants des groupes FW et FB de garder les mains sur les cuisses pendant la phase d’acquisition et de ne pas reproduire les mouvements tout en regardant le(s) modèle(s). La tâche principale de l’expérimentateur était de s’assurer que les participants respectaient ces instructions. Le comportement manifeste des participants suggère qu’ils l’ont fait. Enfin, les participants du groupe témoin ne se sont pas entraînés physiquement ou n’ont rien observé pendant cette phase. Au lieu de cela, ils lisent un journal ou un magazine fourni pendant la même durée que l’observation pour les autres groupes (environ 10 min).
Les troisième et quatrième phases expérimentales étaient des phases de rétention de 10 min et de 24 h. Dans chaque phase, tous les participants ont physiquement effectué 20 essais sans KR. Les participants ont été invités à compléter chaque segment de la tâche en 300 ms, pour une TMT de 1200 ms.
Analyse des données
Les données du pré-test et des deux phases de rétention ont été regroupées en blocs de cinq essais. Pour chaque bloc successif de cinq essais (i.e., essais 1-5, 6-10, etc.), nous avons calculé la valeur absolue de l’erreur constante de chaque participant (|CE|, l’erreur constante indique si un participant a dépassé ou dépassé le temps de mouvement total) et de l’erreur variable du temps de mouvement total (variabilité VE ou intra-participant) pour déterminer la précision et la cohérence de TMT, respectivement. Pour les temps intermédiaires, nous avons calculé un RMSE, qui indique dans quelle mesure chaque participant s’est écarté du schéma de synchronisation relatif prescrit dans un seul score. Pour chaque essai,
où ITi représente le temps intermédiaire pour le segment « i » et target représente le temps de déplacement de l’objectif pour chaque segment de la tâche (c’est-à-dire 300 ms).
Comme les données n’étaient pas normalement distribuées (les données RMSE et temporelles sont biaisées positivement), chaque variable dépendante a subi une transformation logarithmique (ln). Les données transformées pour chaque variable dépendante ont été soumises indépendamment à une ANOVA contrastant trois groupes (C, FW et FB) × trois phases (pré-test, rétention de 10 min, rétention de 24 h) × quatre blocs d’essais (1-5, 6-10, 11-15 et 16-20), avec des mesures répétées sur les deux derniers facteurs. Tous les effets principaux significatifs et les effets principaux simples impliquant plus de deux moyens ont été ventilés à l’aide de l’ajustement de Bonferroni. Pour toutes les comparaisons, un effet a été jugé significatif si p < 0,05. Le carré partiel de l’eta (np2) est la taille de l’effet rapportée pour tous les effets significatifs (Cohen, 1988).
Résultats
Temps total de mouvement
L’ANOVA calculée sur |CE| (Figure 2, panneau supérieur) a révélé des effets principaux significatifs pour le groupe de variables, F(2, 87) = 5,04, p = 0,08, np2 = 0,10 et phase, F(2, 174) = 5,16, p = 0,007, np2 = 0,06, ainsi qu’une interaction de phase × groupe significative, F(4, 174) = 4,93, p = 0,001, np2 = 0,10. La ventilation de cette interaction n’a révélé aucune différence significative de groupe dans le pré-test (F < 1). Dans le test de rétention de 10 minutes, F(2, 87) = 10,12, p < 0,001, np2 = 0,19, les comparaisons post-hoc ont révélé que le groupe témoin avait un | CE | significativement plus grand que les groupes FW et FB (p < 0,05 dans les deux cas), qui ne différaient pas significativement l’un de l’autre (p = 0,19). Dans l’essai de rétention de 24 h, F(2, 87) = 4,34, p = 0,016, np2 = 0,09, le groupe FW avait un | CE/ significativement plus petit que le groupe témoin (p = 0,012)1.
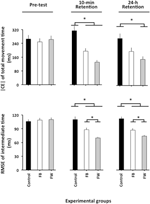
Figure 2. Erreur constante absolue de TMT et erreur quadratique moyenne de la synchronisation relative en fonction des phases expérimentales et des groupes expérimentaux (Expérience 1). * p < 0,05. Les barres d’erreur indiquent l’erreur type de la moyenne.
L’ANOVA calculée sur VE (non représentée) a révélé des effets principaux significatifs pour la phase variable, F(2, 174) = 13,12, p < 0,001, np2 = 0,13, et le bloc, F(3, 261) = 48,79, p < 0,001, np2 = 0,36. Les comparaisons post-hoc de l’effet de phase ont révélé une plus grande VE du temps total dans le pré-test que dans les tests de rétention de 10 min et de 24 h (p < 0.002 dans les deux cas), qui ne différaient pas significativement l’un de l’autre (p = 0,68). L’effet principal du bloc résultait d’une durée totale significativement plus grande pour le premier que pour les trois blocs d’essais restants (p < 0,001 dans tous les cas), qui ne différaient pas significativement les uns des autres (p > 0,05 dans tous les cas).
Synchronisation relative
L’ANOVA calculée sur la RMSE de la synchronisation relative a révélé des effets principaux significatifs pour le groupe de variables, F(2, 87) = 21,49, p < 0,001, np2 = 0,33, phase, F(2, 174) = 39,98, p < 0,001, np2 = 0.31 et bloc, F(3, 261) = 14,77, p < 0,001, np2 = 0,14, ainsi qu’une interaction de phase × groupe significative, F(4, 174) = 12,81, p < 0,001, np2 = 0,23. L’effet principal du bloc résultait d’une RMSE de temps relatif significativement plus importante pour le premier que pour les trois blocs d’essais restants (p < 0,001 dans tous les cas), qui ne différaient pas significativement les uns des autres (p > 0,3 dans tous les cas). Plus intéressant encore, la ventilation de l’interaction phase × groupe (Figure 2, panneau inférieur) n’a révélé aucune différence significative de groupe dans le pré-test (F < 1). Dans les 10 minutes, F(2, 87) = 14,85, p < 0,001, np2 = 0,34, et les tests de rétention sur 24 heures, F(2, 87) = 23,23, p < 0,001, np2 = 0,35, bien que le groupe FB ait nettement surperformé le groupe témoin (p = 0,001 dans les deux cas), le groupe FB a, à son tour, été significativement surperformé par le groupe témoin. Groupe FW (p = 0,001 et p = 0,02, respectivement) 2.
Discussion
La présente expérience a été conçue pour étendre nos connaissances sur les conditions d’observation qui optimisent l’apprentissage d’un nouveau modèle de synchronisation relative. Dans cette situation d’apprentissage, deux groupes d’observation, qui ont observé diverses démonstrations, ont reçu KR avant ou après chaque essai pendant la phase d’acquisition. Plus précisément, nous voulions évaluer si l’apprentissage serait amélioré lorsque les apprenants connaissent la « qualité » ou les caractéristiques d’une démonstration avant d’observer la démonstration. Les résultats sont simples.
Tout d’abord, comme illustré à la figure 2, les groupes FW et FB ont surperformé le groupe témoin lors des tests de rétention. Cela était vrai pour l’apprentissage de la TMT et du timing relatif. Ce résultat attendu confirme les résultats antérieurs qui indiquaient que l’observation permet d’apprendre une nouvelle motricité (voir McCullagh et al., 1989; Hodges et coll., 2007; Vogt et Thomaschke, 2007; Ste-Marie et coll., 2012; Lago-Rodríguez et coll., 2014, pour des revues sur l’apprentissage observationnel) et, notamment, un nouveau modèle de synchronisation relative (Rohbanfard et Proteau, 2011; Andrieux et Proteau, 2013, 2014).
La conclusion la plus importante de la présente étude est que le groupe FB a été surpassé par le groupe FW dans les tests de rétention. Bien que les deux groupes aient observé les mêmes démonstrations, les résultats ont révélé que l’apprentissage est optimisé lorsqu’on connaît à l’avance la qualité ou les caractéristiques de la démonstration témoin. Cette conclusion correspond bien aux rapports précédents de notre laboratoire (Rohbanfard et Proteau, 2011; Andrieux et Proteau, 2013) montrant qu’un régime d’observation mixte, dans lequel les observateurs savent qui est le modèle expert et qui est le modèle novice, favorise l’apprentissage d’un nouveau modèle de synchronisation relative mieux que l’observation experte ou novice seule.
Savoir à l’avance qu’une démonstration moins que parfaite sera démontrée peut être essentiel, étant donné qu’il a été rapporté que les participants novices, comme dans la présente étude, ne sont pas bons pour évaluer la qualité d’une démonstration. Par exemple, Aglioti et al. (2008) ont fait observer aux basketteurs novices et experts des clips vidéo montrant des lancers francs, et les clips vidéo ont été arrêtés à différents moments avant ou immédiatement après la sortie du ballon. Les joueurs de basket-ball experts et les entraîneurs / journalistes spécialisés étaient meilleurs et plus rapides à prédire le sort du tir (réussi ou non) que les novices (pour des résultats similaires, voir aussi Wright et al., 2010; Abreu et coll., 2012; Tomeo et coll., 2013; Balser et coll., 2014; Candidi et coll., 2014; Renden et coll., 2014).
L’avantage du protocole FW par rapport au protocole FB est important et, à notre connaissance, aucune conclusion similaire n’a été rapportée jusqu’à présent. Par conséquent, une réplication de cette constatation semblait importante. De plus, nous nous sommes demandé si l’avantage noté pour le protocole FW ne se produisait qu’après une quantité limitée d’observations. Enfin, nous étions curieux de voir si l’alternance du protocole FW et du protocole FB entraînerait des effets additifs. Nous avons mené l’expérience 2 pour répondre à ces questions.
Expérience 2
Méthodes
Participants
Les 60 participants qui se sont portés volontaires pour cette expérience étaient issus de la même population que celle de l’Expérience 1 (36 hommes et 24 femmes; âge moyen = 22,7 ans; SD = 4,9 ans). Les participants étaient naïfs quant au but de cette étude et n’avaient aucune expérience préalable de la tâche. Ils ont rempli et signé des formulaires de consentement individuels avant de participer. Le Comité d’éthique de la recherche en sciences de la santé de l’Université de Montréal a approuvé cette expérience.
Appareil, Tâche, Phases expérimentales, Procédure et Analyse des données
Nous avons utilisé les mêmes tâches, appareils et procédures que dans l’expérience 1. La différence majeure entre la présente expérience et l’expérience 1 est que les participants ont effectué deux sessions d’acquisition, ce qui a conduit à un total de cinq phases expérimentales: pré-test, acquisition 1, test de rétention immédiate, acquisition 2 et test de rétention 24h.
Les participants ont été assignés au hasard à l’un des trois groupes, chacun composé de 20 participants (8 femelles par groupe): feedforward KR et observation lors de l’acquisition 1 et 2 (FW1-2); feedforward observation et KR lors de l’acquisition 1 mais observation et rétroaction KR lors de l’acquisition 2 (FW/FB); et l’observation et la rétroaction KR lors de l’acquisition 1 et 2 (FB1-2). Nous avons utilisé la même vidéo et les mêmes modèles que dans l’expérience 1; cependant, l’ordre de présentation de la vidéo était différent dans l’acquisition 2 de celui de l’acquisition 1. Tous les participants ont également été informés qu’ils effectueraient la même tâche après chaque phase d’acquisition, mais sans KR concernant leur propre performance.
Nous avons utilisé les mêmes variables dépendantes et la même transformation de données que dans l’expérience 1. Pour chaque variable dépendante, nous avons effectué une ANOVA bidirectionnelle contrastant les trois groupes (FW1-2, FW/FB et FB1-2) × trois phases expérimentales (pré-test, rétention immédiate et rétention 24 h). Tous les effets principaux significatifs et les effets principaux simples impliquant plus de deux moyens ont été ventilés à l’aide de l’ajustement de Bonferroni. Pour toutes les comparaisons, un effet a été jugé significatif si p < 0,05. Le carré partiel de l’eta (np2) est la taille de l’effet rapportée pour tous les effets significatifs (Cohen, 1988).
Résultats
Temps total de mouvement
L’ANOVA calculée pour le |CE| du temps de mouvement (Figure 3) a révélé des effets principaux significatifs pour le groupe de variables, F(2, 57) = 8,13, p = 0,001, np2 = 0,22, et phase, F(2, 114) = 21,13, p < 0,001, np2 = 0,27, ainsi qu’une interaction significative de groupe × phase, F(4, 114) = 2,57, p = 0,042, np2 = 0,08. La ventilation de cette interaction n’a révélé aucune différence significative de groupe dans le pré-test (F < 1). Dans le test de rétention immédiate, F(2, 57) = 10,27, p < 0,002, np2 = 0.27, le groupe FB1-2 avait un | CE | significativement plus grand que les groupes FW1-2 et FW / FB (p < 0,001 dans les deux cas), qui ne différaient pas significativement l’un de l’autre (p > 0,20). Dans l’essai de rétention de 24 h, F(2, 57) = 3,19, p = 0,049, np2 = 0,10, le groupe FW1-2 avait un | CE | légèrement plus petit que le groupe FB1-2 (p = 0,079)3.
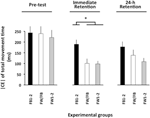
Figure 3. Erreur constante absolue de TMT en fonction des phases expérimentales et des groupes expérimentaux (Expérience 2). * p < 0,05. Les barres d’erreur indiquent l’erreur type de la moyenne.
L’ANOVA calculée sur VE (non représentée) a révélé des effets principaux significatifs pour le groupe de variables, F(2, 57) = 7,82, p = 0,001, np2 = 0,21 et phase, F(2, 114) = 21,10, p < 0,001, np2 = 0,27, ainsi qu’une interaction significative de groupe × phase, F(4, 114) = 4,38, p = 0,002, np2 = 0,13. La ventilation de cette interaction n’a révélé aucune différence de groupe significative dans le pré-test (F < 1) et dans le test de rétention 24 h, F(2, 57) = 1,26, p > 0,20. Dans le test de rétention immédiate, F(2, 57) = 10,26, p < 0,002, np2 = 0,27, le groupe FB1-2 (62.7 ms) avaient une VE significativement plus grande que les groupes FW1-2 (51,1 ms) et FW/FB (53,4 ms) (p < 0,001 dans les deux cas), qui ne différaient pas significativement les uns des autres (p > 0,20)4.
Synchronisation relative
L’ANOVA calculée pour la RMSE de la synchronisation relative a révélé des effets principaux significatifs pour le groupe de variables, F(2, 57) = 4,86, p = 0,01, np2 = 0,15 et phase, F(2, 114) = 78,21, p < 0,001, np2 = 0,58. Il y avait un RMSE de temps relatif significativement plus important dans le pré-test que dans le test de rétention immédiate et le test de rétention 24 h (p < 0.001 dans les deux cas; voir Figure 4, panneau de droite), qui ne différaient pas significativement l’un de l’autre (p > 0,20). Enfin, les groupes FW1-2 et FW/FB ont surperformé le groupe FB1-2 (p = 0,01 et p = 0,07 ; voir Figure 4, panneau de gauche) mais ne différaient pas significativement l’un de l’autre (p > 0,20).
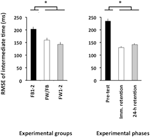
Figure 4. Erreur quadratique moyenne du temps relatif (Expérience 2) en fonction des groupes expérimentaux (panneau de gauche) et des phases expérimentales (panneau de droite). * p < 0,05. Les barres d’erreur indiquent l’erreur type de la moyenne.
Discussion
Comme prévu, la diminution des erreurs constatée lors du passage du pré-test aux tests de rétention corrobore les résultats antérieurs indiquant que l’observation facilite l’apprentissage d’un nouveau modèle de synchronisation relative (Blandin et al., 1999; Rohbanfard et Proteau, 2011; Andrieux et Proteau, 2013, 2014). Plus important encore, les résultats de l’Expérience 2 reproduisaient ceux de l’expérience 1, en ce sens que le groupe FW1-2 surpassait le groupe FB1-2. Par conséquent, on peut conclure sans risque que l’apprentissage de la modification du modèle de synchronisation relative qui émerge naturellement des contraintes de la tâche en un nouveau moment relatif imposé par l’observation est favorisé lorsque l’on est informé des performances du modèle avant plutôt qu’après l’observation. Enfin, les résultats ont également montré que ce qui a été appris dans un protocole FB ne correspond pas à ce qui peut être appris dans un protocole FW.
Discussion générale
L’objectif principal de la présente étude était de déterminer quand, dans un protocole d’observation, KR concernant les performances du modèle devait être fourni, c’est-à-dire avant ou après chaque démonstration. Les résultats des deux expériences de la présente étude ont clairement indiqué que le fait d’être informé des performances du modèle avant chaque démonstration favorisait mieux l’apprentissage d’un nouveau modèle de synchronisation relative que lorsque l’observateur était informé des performances du modèle après chaque démonstration. De plus, les résultats de l’expérience 2 suggèrent que l’avantage du FW par rapport au protocole FB est resté significatif même lorsque le nombre d’essais d’observation a été doublé. Concernant ce dernier point, nous ne soutenons pas qu’un protocole FW devrait être privilégié dans tous les cas et avec tous les niveaux d’expertise des observateurs. Nous soulignons plutôt que l’effet est fiable lorsque l’on considère les observateurs novices.
Nos résultats peuvent indiquer qu’un protocole d’observation prévisionnelle prépare l’observateur à s’engager spécifiquement dans des processus d’imitation lorsqu’une performance experte ou avancée est présentée ou dans des processus de détection d’erreur lorsqu’une performance débutante ou novice est présentée. Cette idée correspond bien aux travaux antérieurs de Decety et al. (1997), qui a déclaré que les schémas d’activation cérébrale pendant l’observation de l’action dépendent à la fois de la nature du traitement exécutif requis et des propriétés extrinsèques de l’action présentée. Plus précisément, ces auteurs ont démontré que différentes zones du cerveau deviennent plus actives lorsque l’on observe pour reconnaître, ce qui pourrait être le cas lors de l’observation d’un modèle novice ou d’une performance médiocre ou intermédiaire, et lorsque l’on observe pour imiter, ce qui est susceptible d’être le cas lors de l’observation d’un modèle expert.
Une autre explication de nos résultats pourrait être qu’un protocole FW entraîne une « désactivation » de l’AON lorsque les participants ont été explicitement informés qu’une mauvaise démonstration suivrait. Par exemple, dans une tâche de levage d’objet, il a été montré que la modulation du potentiel évoqué moteur (MEP) par stimulation magnétique transcrânienne (TMS) lors de l’observation de l’action de levage est mise à l’échelle à la force requise pour effectuer l’action de préhension et de levage (Alaerts et al., 2010a). Il a également été montré que lorsque des indices visuels suggéraient que l’objet était plus lourd qu’il ne l’était réellement, la modulation MEP dépendait principalement du profil cinématique observé plutôt que du poids apparent de l’objet (Alaerts et al., 2010b; Senot et coll., 2011). Cependant, dans une étude de Senot et al. (2011), de fausses informations explicites concernant le poids de l’objet ont été fournies dans une condition expérimentale. Cela a entraîné un conflit entre le profil cinématique attendu compte tenu du poids annoncé et le profil cinématique réel de l’action de préhension et de levage, conduisant à une « inhibition générale du système corticospinal. »Autrement dit, au moins une partie de l’AON avait été désactivée. Par conséquent, il se pourrait que les participants à notre étude aient désactivé l’AON lorsque de mauvaises performances du modèle étaient attendues, laissant l’AON actif uniquement pour de bons essais.
Cette proposition est cependant difficile à concilier avec des rapports récents de notre laboratoire montrant que l’observation à la fois d’un modèle expert et d’un modèle novice a permis de mieux apprendre un nouveau modèle de synchronisation relative que l’observation d’un modèle novice ou d’un modèle expert seul. Si l’on pouvait désactiver l’AON lorsqu’on l’informait qu’une mauvaise démonstration sera présentée (c’est-à-dire un modèle novice), alors l’apprentissage du groupe d’observation mixte aurait égalé et non dépassé celui du groupe d’observation expert. Au contraire, pour en revenir à notre première proposition, nous suggérons qu’un protocole FW aide les artistes débutants à détecter et à quantifier les erreurs dans les performances du modèle, ce qu’ils font généralement mal (Aglioti et al., 2008; Wright et coll., 2010; Abreu et coll., 2012; Tomeo et coll., 2013; Balser et coll., 2014; Candidi et coll., 2014; Renden et coll., 2014). À son tour, la meilleure détection et quantification des performances du modèle peut favoriser le développement de modèles inverses (Jordan, 1996) et forward (Wolpert et Miall, 1996) de contrôle moteur.
En conclusion, l’observation est un outil d’apprentissage puissant qui est accessible à tous et ne nécessite qu’un équipement minimal pour être utilisé. Il est maintenant bien démontré que les avantages de l’observation pour modifier le timing relatif (c’est-à-dire le tempo) de la motricité sont améliorés lorsque l’on a accès à une variété de performances allant des novices aux experts en passant par des horaires d’observation variables ou mixtes. Les résultats de la présente étude suggèrent que ces avantages sont optimisés si l’observateur connaît au préalable la qualité de la performance qu’il s’apprête à observer lors de la première séance d’observation. Cela pourrait être très important dans un contexte de classe dans lequel un enseignant / formateur utiliserait un protocole d’observation vidéo. Par exemple, si l’intention de l’observateur est d’apprendre un aspect spécifique d’un swing de golf, il est probable que le résultat du swing (c’est-à-dire le vol de la balle) ne sera pas montré sur la vidéo. Par conséquent, l’observateur ne serait pas en mesure de « deviner » l’expertise du modèle à partir du résultat du swing et, comme nous l’avons montré dans la présente étude, d’apprendre mieux s’il était informé à l’avance de la qualité de ce qu’il s’apprêtait à observer.
Contributions des auteurs
Tous les auteurs répertoriés ont apporté une contribution substantielle, directe et intellectuelle à l’œuvre et l’ont approuvée pour publication.
Financement
Ce travail a été soutenu par une Subvention à la découverte (LP) fournie par le Conseil de recherches en sciences naturelles et en génie du Canada (subvention no. 111280-2013).
Déclaration de conflit d’intérêts
Les auteurs déclarent que la recherche a été menée en l’absence de relations commerciales ou financières pouvant être interprétées comme un conflit d’intérêts potentiel.
Notes de bas de page
1. ^ Pour vérifier que les différences notées dans les deux tests de rétention entre le groupe témoin et les groupes FW et FB résultaient d’une diminution significative du | CE | du temps de mouvement total, dans une analyse supplémentaire, nous avons décomposé l’interaction groupe × phase rapportée dans le texte principal en calculant une ANOVA distincte pour chaque groupe. Les résultats ont révélé que pour les groupes témoin et FB, le |CE| du temps de mouvement total ne différait pas significativement d’une phase à l’autre, respectivement. Au contraire, pour les groupes FW, il y avait un effet principal significatif des phases, F(2, 86) = 11.60, p < 0,01, np2 = 0,1, qui a révélé une diminution significative du |CE/ du temps de déplacement total allant du pré-test aux deux tests de rétention (p < 0,01), qui ne différaient pas significativement l’un de l’autre (p > 0,10).
2. ^ Comme nous l’avons fait pour le | CE / du temps de mouvement total, dans une analyse supplémentaire, nous avons décomposé l’interaction groupe × phase rapportée dans le texte principal pour le RMSE de la synchronisation relative en calculant une ANOVA distincte pour chaque groupe. Les résultats ont révélé que pour le groupe témoin, le RMSE du timing relatif ne différait pas significativement entre les phases, F(2, 86) = 0,32, p = 0,72, np2 = 0,01. Au contraire, pour les groupes FB et FW, il y a eu un effet principal significatif des phases qui a révélé une diminution significative de la RMSE du timing relatif allant du pré-test aux deux tests de rétention (p < 0,01), qui ne différaient pas significativement l’un de l’autre (p > 0,10).
3. ^ Comme nous l’avons fait dans l’expérience 1, dans une analyse supplémentaire, nous avons décomposé l’interaction groupe × phase rapportée dans le texte principal en calculant une ANOVA distincte pour chaque groupe. Les résultats ont révélé que pour le groupe FB1-2, le / CE/ du temps de mouvement total ne différait pas significativement entre les phases, F(2, 56) < 1, p = 0,45, np2 = 0,03. Au contraire, pour les deux groupes FW1-2 et FW-FB, il y a eu un effet principal significatif des phases, qui, pour les deux groupes, a révélé une diminution significative du |CE/ du temps de déplacement total allant du pré-test aux deux tests de rétention (p < 0.01), qui ne différaient pas significativement les uns des autres (p > 0,10).
4. ^ Pour la VE du temps total de mouvement, la décomposition de l’interaction groupe × phase a révélé un effet principal significatif des phases pour les trois groupes. Pour les groupes FW1-2 et FB1-2, les comparaisons pos-hoc ont révélé un VE significativement plus important dans le pré-test que dans les deux tests de rétention (p < 0,01), qui ne différaient pas significativement l’un de l’autre (p > 0,30). Pour le groupe FW-FB, le VE du temps de mouvement total était significativement plus important dans le pré-test que dans le test de rétention de 24 h (p < 0,01).
Abreu, A. M., Macaluso, E., et Azevedo, R. T. (2012). Anticipation de l’action au-delà du réseau d’observation de l’action: une étude d’imagerie par résonance magnétique fonctionnelle chez des basketteurs experts. EUR. J. Neurosci. 35, 1646–1654. doi: 10.1111 / j.1460-9568.2012.08104.x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Aglioti, S. M., Cesari, P., Romani, M. et Urgesi, C. (2008). Anticipation de l’action et résonance motrice chez les basketteurs d’élite. NAT. Neurosci. 11, 1109–1116. doi: 10.1038/nn.2182
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Al-Abood, S.A., Davids, K. F. et Bennett, S. J. (2001). Spécificité des contraintes de tâche et des effets des démonstrations visuelles et des instructions verbales pour orienter la recherche des apprenants lors de l’acquisition des compétences. J. Mot. Se comporter. 33, 295–305. doi: 10.1080/00222890109601915
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Alaerts, K., Senot, P., Swinnen, S. P., Craighero, L., Wenderoth, N. et Fadiga, L. (2010a). Les exigences de force de levage d’objets observés sont codées par le système moteur de l’observateur: une étude TMS. EUR. J. Neurosci. 31, 1144–1153. doi: 10.1111 / j.1460-9568.2010.07124.x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Alaerts, K., Swinnen, S. P. et Wenderoth, N. (2010b). Observer comment les autres soulèvent des objets légers ou lourds: quels indices visuels interviennent dans le codage de la force musculaire dans le cortex moteur primaire? Neuropsychologie 48, 2082-2090. doi: 10.1016 / j. neuropsychologie.2010.03.029
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Andrieux, M., et Proteau, L. (2013). Observation apprentissage d’une tâche motrice : qui et quand? Exp. Cerveau Res. 229, 125-137. doi: 10.1007/s00221-013-3598- x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Andrieux, M., et Proteau, L. (2014). L’observation mixte favorise l’apprentissage moteur grâce à une meilleure estimation des performances du modèle. Exp. Brain Res. 232, 3121-3132. doi: 10.1007 / s00221-014-4000-3
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Badets, A., et Blandin, Y. (2004). Le rôle de la connaissance de la fréquence des résultats dans l’apprentissage par l’observation. J. Mot. Se comporter. 36, 62–70. doi: 10.3200 / JMBR.36.1.62-70
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Badets, A., et Blandin, Y. (2005). Apprentissage observationnel: effets de la connaissance de la bande passante des résultats. J. Mot. Se comporter. 37, 211–216. doi: 10.3200 / JMBR.37.3.211-216
PubMed Abstract | CrossRef Full Text | Google Scholar
Badets, A., Blandin, Y., Wright, D. L., and Shea, C. H. (2006). Error detection processes during observational learning. Res. Q. Exerc. Sport 77, 177–184. doi: 10.1080/02701367.2006.10599352
PubMed Abstract | CrossRef Full Text | Google Scholar
Balser, N., Lorey, B., Pilgramm, S., Naumann, T., Kindermann, S., Stark, R., et al. (2014). L’influence de l’expertise sur l’activation cérébrale du réseau d’observation d’action lors de l’anticipation du tennis et du volleyball sert. Devant. Hum. Neurosci. 8:568. doi: 10.3389/ fnhum.2014.00568
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Bird, G., et Heyes, C. (2005). Apprentissage dépendant des effecteurs par observation d’une séquence de mouvement des doigts. J. Exp. Psychol. Hum. Percept. Effectuer. 31, 262–275. doi: 10.1037/0096-1523.31.2.262
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Noir, C. B., and Wright, D. L. (2000). Can observational practice facilitate error recognition and movement production? Res. Q. Exerc. Sport 71, 331–339. doi: 10.1080/02701367.2000.10608916
PubMed Abstract | CrossRef Full Text | Google Scholar
Blandin, Y. (1994). Processus Cognitifs Impliqués Lors de L’apprentissage de Tâches de Synchronisation Spatio-Temporelle Sous Différentes Conditions de Pratique et D’observation D’un Sujet Modèle, Thèse De Doctorat Inédite. , Université de Montréal.
Blandin, Y., Lhuisset, L., and Proteau, L. (1999). Processus cognitifs sous-jacents à l’apprentissage observationnel de la motricité. Q. J. Exp. Psychol. 52, 957–979. doi: 10.1080/713755856
CrossRef Texte intégral / Google Scholar
Blandin, Y., Proteau, L. et Alain, C. (1994). Sur les processus cognitifs sous-jacents à l’interférence contextuelle et à l’apprentissage par observation. J. Mot. Se comporter. 26, 18–26. doi: 10.1080/00222895.1994.9941657
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., Gallese, V., et al. (2001). L’observation de l’action active les zones prémotrices et pariétales de manière somatotopique: une étude IRMf. EUR. J. Neurosci. 13, 400–404. doi: 10.1046 /j.1460-9568.2001.01385.x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Buchanan, J.J., et Dean, N. J. (2010). La spécificité dans la pratique profite à l’apprentissage dans les modèles novices et la variabilité dans la démonstration profite à la pratique observationnelle. Psychol. Rés. 74, 313 à 326. doi: 10.1007/s00426-009-0254- y
Résumé PubMed / Texte intégral Croisé / Google Scholar
Buchanan, J. J., Ryu, Y.U., Zihlman, K., et Wright, D.L. (2008). La pratique observationnelle des caractéristiques de mouvement relatif mais non absolu dans une tâche de coordination multi-articulaire à un seul membre. Exp. Brain Res. 191, 157-169. doi: 10.1007/s00221-008-1512-8
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Candidi, M., Sacheli, L. M., Mega, I. et Aglioti, S. M. (2014). Cartographie somatotopique des erreurs de doigté du piano chez des experts sensorimoteurs: Études TMS chez des pianistes et des naïfs musicaux formés visuellement. Céreb. Cortex 24, 435-443. doi: 10.1093/cercor/bhs325
Résumé PubMed / Texte intégral Croisé / Google Scholar
Carroll, W. R. et Bandura, A. (1990). Guide de représentation de la production d’action dans l’apprentissage observationnel: une analyse causale. J. Mot. Se comporter. 22, 85–97. doi: 10.1080/00222895.1990.10735503
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Cisek, P., et Kalaska, J. F. (2004). Corrélats neuraux de répétition mentale dans le cortex prémoteur dorsal. Nature 431, 993-996. doi: 10.1038 / nature03005
Résumé PubMed / Texte intégral Croisé / Google Scholar
Cohen, J. (1988). Analyse de Puissance Statistique pour les Sciences du Comportement. Il s’agit de l’un des plus grands noms de la littérature.
Google Scholar
Collier, G. L., et Wright, C.E. (1995). Redimensionnement temporel de l’échantillon et des rations complexes en tapotement rythmique. J. Exp. Psychol. Hum. Percept. Effectuer. 21, 602–627. doi: 10.1037/0096-1523.21.3.602
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Cross, E. S., Kraemer, D. J. M., Hamilton, A. F. D., Kelley, W. M. et Grafton, S. T. (2009). Sensibilité du réseau d’observation d’action à l’apprentissage physique et observationnel. Céreb. Cortex 19, 315-326. doi: 10.1093/cercor/bhn083
Résumé publié / Texte intégral croisé / Google Scholar
Decety, J., Grèzes, J., Costes, N., Perani, D., Jeannerod, M., Procyk, E., et al. (1997). Activité cérébrale lors de l’observation des actions – influence du contenu de l’action et de la stratégie du sujet. Cerveau 120, 1763-1777. doi: 10.1093/brain/120.10.1763
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Dushanova, J., et Donoghue, J. (2010). Neurones dans le cortex moteur primaire engagés pendant l’observation de l’action. EUR. J. Neurosci. 31, 386–398. doi: 10.1111/j.1460-9568.2009.07067.x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Frey, S. H., et Gerry, V. E. (2006). Modulation de l’activité neuronale lors de l’apprentissage observationnel des actions et de leurs ordres séquentiels. J. Neurosci. 26, 13194–13201. doi: 10.1523/ JNEUROSCI.3914-06.2006
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Gallese V., Fadiga L., Fogassi L., Rizzolatti G. (2002). « Représentation de l’action et lobule pariétal inférieur », dans Common Mechanisms in Perception and Action: Attention and Performance, vol. XIX, éd. W. Prinz et B. Hommel (Oxford : Oxford University Press), 247-266.
Grafton, S. T., Fadiga, L., Arbib, M. A., et Rizzolatti, G. (1997). Activation du cortex promoteur lors de l’observation et de la désignation d’outils familiers. Neuroimage 6, 231-236. doi: 10.1006 / nimg.1997.0293
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Hayes, S. J., Elliott, D. et Bennett, S. J. (2010). Les représentations motrices générales sont développées lors de l’action-observation. Exp. Brain Res. 204, 199-206. doi: 10.1007/s00221-010-2303-6
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Heyes, C. M., et Foster, C.L. (2002). Apprentissage moteur par observation: preuve d’une tâche de temps de réaction en série. Q. J. Exp. Psychol. 55, 593–607. doi: 10.1080/02724980143000389
Résumé PubMed / Texte intégral Croisé / Google Scholar
Il s’agit de la première édition de la série. Le rôle de la vidéo dans la facilitation de la perception et de l’action d’un nouveau mouvement de coordination. J. Mot. Se comporter. 35, 247–260. doi: 10.1080/00222890309602138
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Hodges, N. J., Williams, A. M., Hayes, S. J. et Breslin, G. (2007). Qu’est-ce qui est modélisé lors de l’apprentissage par observation? J. Sport Sci. 25, 531–545. doi: 10.1080/02640410600946860
Résumé PubMed / Texte intégral Croisé / Google Scholar
Jordan, M. I. (1996). « Aspects computationnels du contrôle moteur et de l’apprentissage moteur », dans Handbook of Perception and Action: Vol. 2, Motricité, éd. H. Heuer et S. W. Keele (New York, NY: Academic Press), 71-120.
Il s’agit de l’un des principaux organismes de recherche et de développement du Canada. Preuve de neurones miroirs dans le gyrus frontal inférieur humain. J. Neurosci. 29, 10153–10159. doi: 10.1523/ JNEUROSCI.2668-09.2009
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Lago-Rodríguez, A., Cheeran, B. et Koch, G. (2014). Le rôle des neurones miroirs dans l’apprentissage moteur observationnel: une revue intégrative. EUR. J. Mouvement humain. 32, 82–103.
Lee, T. D., Swinnen, S. P., et Serrien, D. J. (1994). Effort cognitif et apprentissage moteur. Quête 46, 328-344. doi: 10.1080/00336297.1994.10484130
CrossRef Texte intégral / Google Scholar
Lee, T.D., et White, M.A. (1990). Influence d’un programme de pratique de modèles non qualifiés sur l’apprentissage moteur par observation. Hum. Mov. Sci. 9, 349–367. doi: 10.1016/0167-9457(90)90008-2
CrossRef Texte intégral / Google Scholar
Martens, R., Burwitz, L. et Zuckerman, J. (1976). Modélisation des effets sur les performances du moteur. Rés. Q. 47, 277-291.
Résumé PubMed / Google Scholar
McCullagh, P., et Caird, J. K. (1990). Modèles corrects et d’apprentissage et utilisation de la connaissance modèle des résultats dans l’acquisition et la rétention d’une compétence motrice. J. Mouvement humain. Goujon. 18, 107–116.
McCullagh, P., et Meyer, K. N. (1997). Apprentissage versus modèles corrects: influence du type de modèle sur l’apprentissage d’un ascenseur squat poids libre. Rés. Q. Exerc. Sport 68, 56-61. doi: 10.1080/02701367.1997.10608866
Résumé PubMed / Texte intégral CrossRef / Google Scholar
McCullagh, P., Weiss, M. R. et Ross, D. (1989). Considérations de modélisation dans l’acquisition et la performance de la motricité: une approche intégrée. Exercer. Sport Sci. Rév. 17, 475-513.
Résumé PubMed / Google Scholar
Oosterhof, N. N., Wiggett, A. J., Diedrichsen, J., Tipper, S. P., et Downing, P.E. (2010). La cartographie de l’information basée sur la surface révèle des représentations vision-action crossmodales dans le cortex pariétal et occipitotemporal humain. J. Neurophysiol. 104, 1077–1089. doi: 10.1152/jn.00326.2010
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Pollock, B. J. et Lee, T.D. (1992). Effets du niveau de compétence du modèle sur l’apprentissage moteur par observation. Rés. Q. Exerc. Sport 63, 25-29. doi: 10.1080/02701367.1992.10607553
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Il s’agit de l’un des plus grands ouvrages de l’histoire de l’art et de l’histoire de l’Art. Faute ou plongée? Contributions motrices au jugement des situations de faute ambiguës dans le football. EUR. J. Sport Sci. 14 (Suppl. 1), S221–S227. doi: 10.1080/17461391.2012.683813
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Rizzolatti, G., Cattaneo, L., Fabbri-Destro, M., et Rozzi, S. (2014). Mécanismes corticaux sous-jacents à l’organisation des actions dirigées vers des objectifs et à la compréhension des actions basées sur les neurones miroirs. Physiol. Rév. 94, 655-706. doi: 10.1152 / physrev.00009.2013
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Rizzolatti, G., et Fogassi, L. (2014). The mirror mechanism: recent findings and perspectives. Philo. Trans. R. Soc. Lond. Ser. B Biol. Sci. 369:20130420. doi: 10.1098/rstb.2013.0420
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Rohbanfard, H., et Proteau, L. (2011). Apprendre par l’observation: une combinaison de modèles experts et novices favorise l’apprentissage. Exp. Brain Res. 215, 183-197. doi: 10.1007/s00221-011-2882- x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Senot, P., D’Ausilio, A., Franca, M., Caselli, L., Craighero, L., et Fadiga, L. (2011). Effect of weight-related labels on corticospinal excitability during observation of grasping: a TMS study. Exp. Cerveau Res. 211, 161-167. doi: 10.1007 / s00221-011-2635- x
Résumé PubMed / Texte intégral Croisé / Google Scholar
Ste-Marie, D. M., Law, B., Rymal, A. M., Jenny, O., Hall, C., et McCullagh, P. (2012). Interventions d’observation pour l’apprentissage de la motricité et la performance: un modèle appliqué pour l’utilisation de l’observation. Int. Rév. Sport Exerc. Psychol. 5, 145–176. doi: 10.1080/1750984X.2012.665076
CrossRef Texte intégral / Google Scholar
Tomeo, E., Cesari, P., Aglioti, S. M., et Urgesi, C. (2013). Tromper les botteurs mais pas les gardiens de but: corrélats comportementaux et neurophysiologiques de la détection de fausses actions dans le football. Céreb. Cortex 23, 2765-2778. doi: 10.1093/cercor/bhs279
Résumé publié / Texte intégral croisé / Google Scholar
Vogt, S., et Thomaschke, R. (2007). Des interactions visuo-motrices à l’apprentissage de l’imitation : études comportementales et d’imagerie cérébrale. J. Sport Sci. 25, 497–517. doi: 10.1080/02640410600946779
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Wolpert, D. M., et Miall, R. C. (1996). Modèles avancés pour le contrôle physiologique du moteur. Réseau neuronal. 9, 1265–1279. doi: 10.1016/S0893-6080(96)00035-4
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Il s’agit de l’un des principaux facteurs de risque de la maladie de Parkinson. Processus cognitifs liés à l’interférence contextuelle et à l’apprentissage observationnel: une réplication de Blandin, Proteau et Alain (1994). Rés. Q. Exerc. Sport 68, 106-109. doi: 10.1080/02701367.1997.10608872
Résumé PubMed / Texte intégral CrossRef / Google Scholar
Le président de la République, M. J., Bishop, D. T., Jackson, R. C., and Abernethy, B. (2010). Functional MRI reveals expert-novice differences during sport-related anticipation. Neuroreport 21, 94–98. doi: 10.1097/WNR.0b013e328333dff2
PubMed Abstract | CrossRef Full Text | Google Scholar