- Introduksjon
- Eksperiment 1
- Metoder
- Deltakere
- Apparater Og Oppgaver
- Eksperimentelle Faser Og Prosedyrer
- Dataanalyse
- Resultater
- Total Bevegelsestid
- Relativ Timing
- Diskusjon
- Eksperiment 2
- Metoder
- Deltakere
- Apparater, Oppgave, Eksperimentelle Faser, Prosedyre Og Dataanalyse
- Resultater
- Total Bevegelsestid
- Relativ Timing
- Diskusjon
- Generell Diskusjon
- Forfatterbidrag
- Finansiering
- Interessekonflikt
- Fotnoter
Introduksjon
Du er en ivrig golfspiller og du vil lære et nytt skudd. Hvordan ville du fortsette? Det er en god sjanse for at du vil observere noen (live, på video, På Youtube, etc.) hvem vet hvordan du skal utføre dette bildet, og du vil prøve å forstå hva du skal gjøre og hvordan du gjør det. Forskning tyder tydelig på at denne læringsstrategien er vellykket fordi observasjon har vist seg å fremme læring av et bredt spekter av motoriske ferdigheter (se McCullagh et al., 1989; Hodges et al., 2007; Vogt Og Thomaschke, 2007; Ste-Marie et al., 2012; Lago-Rodrí Et al., 2014, for vurderinger om observasjonell læring). Dette skyldes at observasjon har mye felles med fysisk praksis, som er den første determinanten av motorisk ferdighetslæring. Spesielt har det blitt vist at variabler, for eksempel mengden praksis (Carroll Og Bandura, 1990; Blandin, 1994), hyppigheten av kunnskap om resultater (, Badets og Blandin, 2004, 2005; Badets et al., 2006), og praksisplanen (Blandin et al., 1994; Wright et al., 1997), påvirker læring via observasjonspraksis og fysisk praksis på lignende måter. Disse dataene førte til forslaget om at observasjon og fysisk praksis bruker svært lignende prosesser. Dette forslaget støttes av resultatene av neuroimaging studier som viste at et ensemble av nevrale strukturer (inkludert premotor cortex, den dårligere parietal lobule, superior temporal sulcus, supplerende motorområdet, cingulate gyrus og cerebellum), også kalt «action observation network» (Aon) (Kilner et al., 2009; Oosterhof et al., 2010), aktiveres både når enkeltpersoner utfører en gitt motoroppgave og når de observerer andre som utfører den samme motoroppgaven (Grafton et al., 1997; Buccino et al., 2001; Gallese et al., 2002; Cisek Og Kalaska, 2004; Frey Og Gerry, 2006; Cross et al., 2009; Dushanova Og Donoghue, 2010; Rizzolatti Og Fogassi, 2014; Rizzolatti et al., 2014).
Observasjon favoriserer motorisk ferdighetslæring, men hvem bør du observere for å lære det nye golfskuddet? En ekspert som mestrer skuddet, vil antagelig hjelpe deg med å utvikle en referanse til hva du skal gjøre og hvordan du gjør det, men bør du observere noen som deg som lærer det skuddet og som antagelig gir deg en bedre sjanse til å oppdage og lære av feil eller endringer i strategi? Forskning har vist at observere både en dyktig modell (Martens et al., 1976; McCullagh et al., 1989; Lee et al., 1994; Al-Abood et al., 2001; Heyes og Foster, 2002; Hodges et al., 2003; Bird Og Heyes, 2005) og en nybegynnermodell fører til betydelig læring (Lee Og White, 1990; McCullagh Og Caird, 1990; Pollock Og Lee, 1992; McCullagh Og Meyer, 1997; Black Og Wright, 2000; Buchanan et al., 2008; Buchanan og Dean, 2010; Hayes et al., 2010). Nylige resultater fra vårt laboratorium viste imidlertid at observasjonslæring av en ny motorisk ferdighet er forbedret etter observasjon av både nybegynnere og ekspertmodeller i stedet for enten en nybegynner eller en ekspertmodell alene (Rohbanfard og Proteau, 2011; Andrieux og Proteau, 2013, 2014). Vi tror at dette» variable » observasjonsformatet fører til ikke bare utvikling av en god bevegelsesrepresentasjon (ekspertobservasjon), men også utvikling av effektive prosesser for feildeteksjon og korreksjon (nybegynnerobservasjon).
i denne studien er spørsmålet om interesse en enkel, men viktig en. Ved bruk av en variabel observasjonsplan, vil læring bli bedre når observatørene blir informert på forhånd om» kvaliteten » på forestillingen de skal se, eller vil det bli bedre når observatørene blir igjen for å evaluere forestillingene før de mottar tilbakemelding. Å informere observatørene om hva de er i ferd med å se, kan gjøre dem i stand til å velge om de vil observere for å etterligne eller heller observere for å oppdage feil eller svakheter i modellens ytelse, noe som kan lette utviklingen av disse prosessene. Alternativt kan deltakerne evaluere ytelseskvaliteten de observerte aktivere mer forseggjorte kognitive prosesser enn når denne informasjonen blir matet fremover (f.eks. feildeteksjon og anerkjennelse eller evaluering av alternativ strategi), noe som resulterer i bedre læring av oppgaven.
oppgaven som vi valgte, krevde deltakerne å endre det relative timemønsteret som naturlig oppsto fra oppgavebegrensningene (Collier Og Wright, 1995; Blandin et al., 1999) til et nytt, pålagt mønster av relativ timing. Dette ligner på å endre tempoet når man utfører en serve i tennis eller en stasjon i golf (Rohbanfard og Proteau, 2011). Deltakerne observert to modeller som demonstrerer et bredt spekter av forestillinger. I en gruppe ble observatører informert før hvert forsøk om kvalitetsnivået (ekspert, avansert, middels, nybegynner eller nybegynner) av hva de var i ferd med å se, mens en annen gruppe observatører ble gitt den samme informasjonen først etter at hver observasjonsstudie var fullført.
Eksperiment 1
Metoder
Deltakere
Nitti høyrehendte studenter (45 menn og 45 kvinner; gjennomsnittsalder = 20,5 år; SD = 0,9 år) Fra Déé deltok i dette eksperimentet. Deltakerne var naive til formålet med studien og hadde ingen tidligere erfaring med oppgaven, og alle deltakerne var selverklærte som høyrehendte. Ingen av deltakerne rapporterte nevrologiske lidelser, og alle hadde normalt eller korrigert til normalt syn. Deltakerne fullførte og signerte individuelle samtykkeskjemaer før deltakelse. Forskningsetiske Komiteer For Helsevitenskap Ved Universiteteté De Montréal godkjente dette eksperimentet.
Apparater Og Oppgaver
apparatet var lik Det Som Ble brukt Av Rohbanfard and Proteau (2011). Som illustrert i Figur 1 besto den av en trebunn (45 × 54 cm), tre trebarrierer (11 × 8 cm) og en startknapp innebygd i et mål (11 × 8 cm). Avstanden mellom startknappen og den første barrieren var 15 cm. Avstandene til de resterende tre segmentene av oppgaven var henholdsvis 32, 18 og 29 cm. Barrierene ble plassert vinkelrett på trebunnen i begynnelsen av hvert forsøk, noe som ga en lukket mikrobryterkrets. Alle mikrobrytere ble koblet til en datamaskin via I / o-porten til En A-d-omformer (National Instruments, Austin, Texas, USA), og en millisekund timer ble brukt til å registrere både total bevegelsestid (TMT) og tiden som kreves for å fullføre hvert segment av oppgaven (mellomtider, ITs).
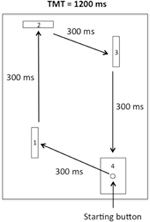
Figur 1. Skisse av apparatet. Deltakerne måtte forlate startknappen og slå den første, andre og tredje barrierene med urviseren før de endelig nådde målet.
for de fysiske praksisforsøkene (se nedenfor) satt deltakerne nær startposisjonen foran apparatet. Deretter, fra startknappen, deltakerne ble bedt om å suksessivt slå ned den første, andre og tredje barrierer (dermed slippe mikrobrytere) og til slutt treffer målet med klokken som illustrert I Figur 1. Hvert segment av oppgaven måtte fullføres I EN IT på 300 ms, for EN TMT på 1200 ms. Bevegelsesmønsteret, Dets OG TMT ble illustrert på en plakat som ligger rett foran apparatet under alle eksperimentelle faser.
Eksperimentelle Faser Og Prosedyrer
deltakerne ble randomisert til en av de tre gruppene, hver bestående av 30 deltakere (15 kvinner per gruppe): kontroll (C), feedforward KR OG observasjon (fw), og observasjon OG tilbakemelding KR (FB). Alle gruppene utførte fire eksperimentelle faser, fordelt over 2 påfølgende dager.
alle deltakerne fikk muntlige instruksjoner om TMT og ITS før første eksperimentelle fase. Den første eksperimentelle fasen var en forprøve, hvor alle deltakerne utførte 20 fysiske praksisforsøk uten kjennskap til resultatene (KR) på TMT og ITs.
den andre fasen var en oppkjøpsfase og besto av 60 observasjonsstudier for deltakerne i de to observasjonsgruppene (FW og FB). Disse deltakerne så individuelt på en videopresentasjon av to modeller som fysisk utførte den eksperimentelle oppgaven. FOR hver observasjonsstudie ble KR om modellens ytelse (BÅDE TMT og ITS) presentert i ms (Se Figur 1) enten før demonstrasjonen for FW-gruppen eller etter demonstrasjonen for FB-gruppen. Modellen ble endret hvert femte forsøk (dvs. modell 1: forsøk 1-5 og modell 2: forsøk 6-10, og så videre), for totalt 30 forsøk utført av en modell og 30 forsøk utført av den andre modellen. For BÅDE FW-og FB-gruppene ble de to modellene, som deltok i tidligere arbeid fra vårt laboratorium, valgt fordi for begge modellene hadde vi seks videoklipp som illustrerte forestillinger i hver av fem underkategorier. Dermed kunne deltakerne I fw-og FB-gruppene ikke knytte en bestemt modell med enten bedre eller dårligere ytelse. En ekspert ytelse samsvarer med en rot gjennomsnittlig kvadrat feil (RMSE; se dataanalyse delen for beregning detaljer) varierer mellom 0 og 15 ms; avansert, middels, nybegynner og nybegynner forestillinger korresponderte Med RMSEs på henholdsvis 30-45 ms, 60-75 ms, 90-105 ms og 120+ ms. Deltakerne i FW-og FB-gruppene ble informert om modellens ytelse i ms; de ble også informert om ytelsesnivået som det refererte til. De resulterende 30 forsøkene av hver modell (fem nivåer av ytelse × seks repetisjoner) ble randomisert slik at de fem nivåene av ytelse ble presentert en gang i hvert sett med fem forsøk. For å unngå fysisk etterligning av sekvensen, som kan forstyrre observasjonsprosessene, spurte vi deltakerne I FW-og FB-gruppene om å holde hendene på lårene i oppkjøpsfasen og ikke reprodusere bevegelsene mens de så på modellen(e). Det Var Eksperimentatorens hovedoppgave å sikre at deltakerne fulgte disse instruksjonene. Deltakernes åpenbare oppførsel antyder at de gjorde det. Til slutt trente deltakerne i kontrollgruppen ikke fysisk eller observert noe i denne fasen. I stedet leser de en gitt avis eller et magasin i samme varighet som observasjonen for de andre gruppene(omtrent 10 min).
den tredje og fjerde eksperimentelle fasen var 10-min og 24-h retensjonsfaser. I hver fase utførte alle deltakerne fysisk 20 forsøk uten KR. Deltakerne ble bedt om å fullføre hvert segment av oppgaven i 300 ms, for EN TMT på 1200 ms.
Dataanalyse
dataene fra pre-testen og de to retensjonsfasene ble omgruppert i blokker med fem forsøk. For hver påfølgende blokk med fem forsøk (dvs., forsøk 1-5, 6-10, etc.), beregnet vi absoluttverdien av hver deltakers konstante feil (|CE|, den konstante feilen indikerer om en deltaker under eller overshot den totale bevegelsestiden) og variabel feil av den totale bevegelsestiden (VE eller innenfor deltakerens variabilitet) for å bestemme nøyaktigheten og konsistensen av TMT, henholdsvis. For mellomtider, vi beregnet EN RMSE, som angir hvor mye hver deltaker avvek fra den foreskrevne relative timing mønster i en enkelt poengsum. FOR hvert forsøk,
Hvor ITi representerer mellomtiden for segmentet «i», og målet representerer målbevegelsestiden for hvert segment av oppgaven (dvs. 300 ms).
fordi dataene ikke var normalfordelt (RMSE og tidsdata er positivt skjev), gjennomgikk hver avhengig variabel en logaritmisk transformasjon (ln). De transformerte dataene for hver avhengig variabel ble uavhengig sendt til EN ANOVA som kontrasterte tre grupper (C, FW og FB) × tre faser (pre-test, 10-min retensjon, 24-h retensjon) × fire blokker av forsøk (1-5, 6-10, 11-15 og 16-20), med gjentatte tiltak på de to siste faktorene. Alle de betydelige hovedeffekter og enkle hovedeffekter som involverer mer enn to midler ble brutt ned Ved Hjelp Av Bonferroni justering. For alle sammenligninger ble en effekt ansett som signifikant hvis p < 0,05. Delvis eta-kvadrat (np2) er effektstørrelsen rapportert for alle signifikante effekter (Cohen, 1988).
Resultater
Total Bevegelsestid
ANOVA beregnet på |CE| (Figur 2, øvre panel) viste signifikante hovedeffekter for variabelgruppen, F(2, 87) = 5,04, p = 0,08, np2 = 0,10 og fase, F(2, 174) = 5,16, p = 0,007, np2 = 0,06, som i tillegg til en signifikant fase × gruppeinteraksjon, f(4, 174) = 4,93, P = 0,001, Np2 = 0,10. Fordelingen av denne interaksjonen viste ingen signifikante gruppeforskjeller i forprøven (F < 1). I 10-min retensjonstest, F(2, 87) = 10,12, p < 0,001, np2 = 0,19, viste post-hoc-sammenligningene at kontrollgruppen hadde en signifikant større | CE| enn BÅDE FW-og FB-gruppene (p < 0,05 i begge tilfeller), som ikke var signifikant forskjellig fra hverandre (p = 0,19). I 24-h retensjonstesten, F(2, 87) = 4,34, p = 0,016, np2 = 0,09, HADDE FW-gruppen en betydelig mindre / CE / enn kontrollgruppen(p = 0,012) 1.
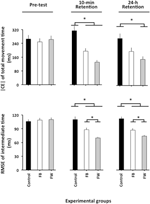
Figur 2. Absolutt konstant FEIL AV TMT og rot betyr kvadratfeil av relativ timing som en funksjon av eksperimentelle faser og eksperimentelle grupper (Eksperiment 1). * p < 0,05. Feilfelt angir standardfeil av gjennomsnittet.
ANOVA beregnet PÅ VE (ikke vist) viste signifikante hovedeffekter for den variable fasen, F(2, 174) = 13,12, p < 0,001, np2 = 0,13 og blokk, F(3, 261) = 48,79, p < 0,001, np2 = 0,36. Post-hoc-sammenligninger av faseeffekten viste en STØRRE VE av total tid i pre-testen enn i både 10-min og 24-h retensjonstestene (p < 0.002 i begge tilfeller), som ikke skilte seg vesentlig fra hverandre (p = 0,68). Blokkhovedeffekten skyldtes en signifikant større VE av total tid for den første enn for de tre gjenværende blokkene av forsøk (p < 0,001 i alle tilfeller), som ikke skilte seg vesentlig fra hverandre(p > 0,05 i alle tilfeller).
Relativ Timing
ANOVA beregnet PÅ RMSE av relativ timing viste signifikante hovedeffekter for variabelgruppen, F(2, 87) = 21,49, p < 0,001, np2 = 0,33, fase, F(2, 174) = 39,98, p < 0,001, np2 = 0.31 og blokk, F(3, 261) = 14,77, p < 0,001, np2 = 0,14, Samt en signifikant fase × gruppeinteraksjon, F(4, 174) = 12,81, p < 0,001, np2 = 0,23. Blokkens hovedeffekt skyldtes en signifikant større RMSE av relativ timing for den første enn for de tre gjenværende blokkene av forsøk (p < 0,001 i alle tilfeller), som ikke skilte seg vesentlig fra hverandre(p> 0,3 i alle tilfeller). Mer interessant var det at fordelingen av fase × gruppeinteraksjonen (Figur 2, nedre panel) ikke viste noen signifikante gruppeforskjeller i forprøven (F < 1). I 10-min, F(2, 87) = 14,85, p < 0,001, np2 = 0,34 og 24-h retensjonstestene, F(2, 87) = 23,23, p < 0,001, np2 = 0,35, selv om FB-gruppen signifikant overgikk kontrollgruppen (p = 0,001 i begge tilfeller), VAR FB-gruppen i sin tur betydelig overgått av fw-gruppen (p = 0,001 og p = 0,02, henholdsvis)2.
Diskusjon
det nåværende eksperimentet ble designet for å utvide vår kunnskap om observasjonsforholdene som optimaliserer læring av et nytt relativt timemønster. I denne læringssituasjonen ble to observasjonsgrupper, som observerte en rekke demonstrasjoner, gitt KR enten før eller etter hvert forsøk i oppkjøpsfasen. Spesielt ønsket vi å vurdere om læring ville bli forbedret når elevene kjenner» kvaliteten » eller egenskapene til en demonstrasjon før de observerer demonstrasjonen. Resultatene er enkle.
først, som illustrert i Figur 2, utkonkurrerte BÅDE FW-og FB-gruppene kontrollgruppen på retensjonstestene. Dette var sant for læring av BÅDE TMT og den relative timingen. Dette forventede resultatet bekrefter tidligere funn som indikerte at observasjon gjør det mulig å lære en ny motorisk ferdighet (se McCullagh et al., 1989; Hodges et al., 2007; Vogt Og Thomaschke, 2007; Ste-Marie et al., 2012; Lago-Rodrí Et al., 2014, for vurderinger om observasjonslæring) og spesielt et nytt relativt timingsmønster (Rohbanfard and Proteau, 2011; Andrieux and Proteau, 2013, 2014).
det viktigste funnet i denne studien er AT FB-gruppen ble overgått AV FW-gruppen i retensjonstestene. Selv om de to gruppene observerte de samme demonstrasjonene, viste resultatene at læring er optimalisert når man får forhåndskunnskap om kvaliteten eller egenskapene til den vitne demonstrasjonen. Dette funnet passer godt med tidligere rapporter fra vårt laboratorium (Rohbanfard og Proteau, 2011; Andrieux og Proteau, 2013) som viser at et blandet observasjonsregime, der observatørene vet hvem som er ekspertmodellen og hvem som er nybegynnermodellen, favoriserer læring av et nytt relativt timingsmønster bedre enn enten ekspert eller nybegynner observasjon alene.
å ha forhåndskunnskap om at en mindre enn perfekt demonstrasjon vil bli vist, kan være kritisk, med tanke på at det har blitt rapportert at nybegynnere, som i den nåværende studien, ikke er gode til å vurdere kvaliteten på en demonstrasjon. For eksempel, Aglioti et al. (2008) hadde nybegynnere og ekspert basketballspillere observere videoklipp som viser frikastskudd, og videoklippene ble stoppet på forskjellige tidspunkter før eller umiddelbart etter ballutgivelsen. Ekspert basketballspillere og trenere/spesialiserte journalister var bedre og raskere til å forutsi skjebnen til skuddet (vellykket eller ikke) enn nybegynnere (for lignende resultater se Også Wright et al., 2010; Ove et al., 2012; Tomeo et al., 2013; Balser et al., 2014; Kristian et al., 2014; Renden et al., 2014).
fordelen MED FW over FB-protokollen er viktig, og så vidt vi vet har det ikke blitt rapportert et lignende funn hittil. Derfor virket en replikering av dette funnet viktig. I tillegg lurte vi på om fordelen notert FOR FW-protokollen bare skjedde etter en begrenset mengde observasjon. Til slutt var vi nysgjerrige på å se om alternerende FW og FB-protokollen ville resultere i additive effekter. Vi gjennomførte Eksperiment 2 for å løse disse spørsmålene.
Eksperiment 2
Metoder
Deltakere
de 60 deltakerne som meldte seg frivillig til dette eksperimentet ble hentet fra Samme populasjon som Eksperiment 1 (36 menn og 24 kvinner; gjennomsnittsalder = 22,7 år; SD = 4,9 år). Deltakerne var naive om formålet med denne studien og hadde ingen tidligere erfaring med oppgaven. De fullførte og signerte individuelle samtykkeskjemaer før deltakelse. Forskningsetiske Komiteer For Helsevitenskap Ved Universiteteté De Montréal godkjente dette eksperimentet.
Apparater, Oppgave, Eksperimentelle Faser, Prosedyre Og Dataanalyse
vi brukte samme oppgave, apparat og prosedyrer som I Eksperiment 1. Den store forskjellen mellom det nåværende eksperimentet og Eksperiment 1 er at deltakerne utførte to oppkjøpsøkter, noe som førte til totalt fem eksperimentelle faser: pre-test, oppkjøp 1, umiddelbar retensjonstest, oppkjøp 2 og 24-h retensjonstest.
deltakerne ble randomisert til en av de tre gruppene, hver bestående av 20 deltakere( 8 kvinner per gruppe): feedforward KR OG observasjon under både oppkjøp 1 og 2 (FW1-2); feedforward observasjon og KR under oppkjøp 1 men observasjon OG tilbakemelding KR under oppkjøp 2 (FW / FB); og observasjon og KR tilbakemelding under både oppkjøp 1 og 2 (FB1-2). Vi brukte samme video og modeller som I Eksperiment 1; men rekkefølgen på videopresentasjon var forskjellig i oppkjøp 2 fra det i oppkjøp 1. Alle deltakerne ble også informert om at de ville utføre samme oppgave etter hver oppkjøpsfase, men uten KR om egen ytelse.
vi brukte de samme avhengige variablene og datatransformasjonen som I Eksperiment 1. For hver avhengig variabel gjennomførte vi en toveis ANOVA som kontrasterte de tre gruppene (FW1-2, FW/FB og FB1-2) × tre eksperimentelle faser (pre-test, umiddelbar retensjon og 24-h retensjon). Alle de betydelige hovedeffekter og enkle hovedeffekter som involverer mer enn to midler ble brutt ned Ved Hjelp Av Bonferroni justering. For alle sammenligninger ble en effekt ansett som signifikant hvis p < 0,05. Delvis eta-kvadrat (np2) er effektstørrelsen rapportert for alle signifikante effekter (Cohen, 1988).
Resultater
Total Bevegelsestid
ANOVA beregnet for |CE| av bevegelsestid (Figur 3) viste signifikante hovedeffekter for variabelgruppen, F(2, 57) = 8,13, p = 0,001, np2 = 0,22 og fase, F(2, 114) = 21,13, p < 0,001, np2 = 0,27, Samt En Signifikant Gruppe × Faseinteraksjon, F(4, 114) = 2,57, P = 0,042, Np2 = 0,08. Fordelingen av denne interaksjonen viste ingen signifikante gruppeforskjeller i forprøven (F < 1). I den umiddelbare retensjonstesten, F(2, 57) = 10,27, p < 0,002, np2 = 0.27, FB1 – 2-gruppen hadde en betydelig større | CE| enn BÅDE FW1-2 og FW/FB-gruppene (p < 0,001 i begge tilfeller), som ikke skilte seg vesentlig fra hverandre (p>0,20). I 24-h retensjonstesten, F(2, 57) = 3,19, p = 0,049, np2 = 0,10, HADDE FW1-2-gruppen en litt mindre / CE / enn FB1 – 2-gruppen (p = 0,079)3.
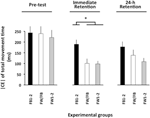
Figur 3. Absolutt konstant FEIL AV TMT som en funksjon av eksperimentelle faser og eksperimentelle grupper (Eksperiment 2). * p < 0,05. Feilfelt angir standardfeil av gjennomsnittet.
ANOVA beregnet PÅ VE (ikke vist) viste signifikante hovedeffekter for variabelgruppen, F(2, 57) = 7,82, p = 0,001, np2 = 0,21, og fase, F(2, 114) = 21,10, p < 0,001, np2 = 0,27, samt en signifikant gruppe × faseinteraksjon, F(4, 114) = 4,38, p = 0,002, np2 = 0,13. Fordelingen av denne interaksjonen viste ingen signifikante gruppeforskjeller i forprøven (F < 1) og i 24-h retensjonstesten, F(2, 57) = 1,26, p> 0,20. I den umiddelbare retensjonstesten, F(2, 57) = 10,26, p < 0,002, np2 = 0,27, fb1-2-gruppen (62.7 ms) hadde en signifikant større VE enn både FW1-2 (51,1 ms) og fw/FB (53,4 ms) gruppene (p < 0,001 i begge tilfeller), som ikke skilte seg vesentlig fra hverandre (p>0,20)4.
Relativ Timing
ANOVA beregnet FOR RMSE av relativ timing viste signifikante hovedeffekter for variabelgruppen, F(2, 57) = 4,86, p = 0,01, np2 = 0,15 og fase, F(2, 114) = 78,21, p < 0,001, np2 = 0,58. DET var en signifikant større RMSE av relativ timing i forprøven enn i både umiddelbar retensjonstest og 24-h retensjonstest (p < 0.001 i begge tilfeller, se Figur 4, høyre panel), som ikke skilte seg vesentlig fra hverandre (p>0,20). TIL slutt overgikk fw1-2-og fw/FB-gruppene FB1-2-gruppen (p = 0,01 og p = 0,07; se Figur 4, venstre panel), men var ikke signifikant forskjellig fra hverandre (p>0,20).
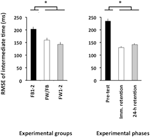
Figur 4. Root mean square feil av relativ timing (Eksperiment 2) som en funksjon av eksperimentelle grupper (venstre panel) og eksperimentelle faser (høyre panel). * p < 0,05. Feilfelt angir standardfeil av gjennomsnittet.
Diskusjon
som forventet, nedgangen i feil bemerket når du går fra pre-test til oppbevaring tester støtter tidligere funn som indikerer at observasjon hjelpemidler læring av en ny relativ timing mønster (Blandin et al.( 1999; Rohbanfard Og Proteau, 2011; Andrieux Og Proteau, 2013, 2014). Enda viktigere, resultatene Av Eksperiment 2 replikerte De Av Eksperiment 1, ved AT fw1-2-gruppen overgikk FB1 – 2-gruppen. Derfor kan det trygt konkluderes med at læring for å endre det relative timingmønsteret som naturlig kommer fra oppgavens begrensninger til en ny, pålagt relativ timing gjennom observasjon, favoriseres når man blir informert om modellens ytelse før i stedet for etter observasjon. Til slutt viste resultatene også at det som har blitt lært I EN FB-protokoll, ikke legger opp til det som kan læres i EN FW-protokoll.
Generell Diskusjon
hovedmålet med denne studien var å bestemme når I en observasjonsprotokoll SKAL KR om modellens ytelse gis, dvs. før eller etter hver demonstrasjon. Resultatene fra de to forsøkene i denne studien viste tydelig at det å bli informert om modellens ytelse før hver demonstrasjon favoriserte læring av et nytt relativt timingsmønster bedre enn når observatøren ble informert om modellens ytelse etter hver demonstrasjon. Videre tyder resultatene av Eksperiment 2 på at fordelen AV FW over FB-protokollen forblir signifikant selv når antall observasjonsforsøk ble doblet. Når det gjelder dette siste punktet, argumenterer vi ikke for at EN FW-protokoll bør favoriseres i alle tilfeller og med alle nivåer av ekspertise fra observatørene. Snarere understreker vi at effekten er pålitelig når nybegynnere observatører vurderes.
våre resultater kan indikere at en feedforward observasjonsprotokoll forbereder observatøren til å engasjere seg spesifikt i enten imitasjonsprosesser når en ekspert eller avansert ytelse vises eller i feildeteksjonsprosesser når en nybegynner eller nybegynner ytelse presenteres. Denne ideen passer godt med tidligere arbeid Fra Decety et al. (1997), som uttalte at mønstrene av hjerneaktivering under handlingsobservasjon avhenger av både arten av den nødvendige utøvende behandlingen og de ytre egenskapene til handlingen som presenteres. Spesielt viste disse forfatterne at ulike områder av hjernen blir mer aktive når man observerer å gjenkjenne, noe som kan være tilfelle når man observerer en nybegynnermodell eller en dårlig eller mellomliggende ytelse, og når man observerer å etterligne, noe som sannsynligvis vil være tilfelle når man observerer en ekspertmodell.
en alternativ forklaring på våre funn kan være at EN FW-protokoll resulterer i en» deaktivering » av AON når deltakerne ble eksplisitt informert om at en dårlig demonstrasjon ville følge. For eksempel, i en objektløfteoppgave, har det blitt vist at moduleringen av motorisk fremkalt potensial (MEP) ved transkraniell magnetisk stimulering (TMS) under observasjon av løftevirkningen skaleres til kraften som kreves for å utføre grep og løftevirkningen (Alaerts et al.( 2010a). Det ble også vist at når visuelle signaler antydet at objektet var tyngre enn i egentlig var, mep modulering avhengig primært på den observerte kinematiske profil snarere enn på den tilsynelatende vekten av objektet (Alaerts et al., 2010b; Senot et al., 2011). Men i en studie Av Senot et al. (2011), falsk eksplisitt informasjon om vekten av objektet ble gitt i en eksperimentell tilstand. Dette resulterte i en konflikt mellom den forventede kinematiske profilen gitt den annonserte vekten og den faktiske kinematiske profilen til gripe – og løfteaksjonen, noe som førte til en » generell hemming av kortikospinalsystemet.»Uttalt annerledes, minst en del AV AON hadde blitt slått av. Derfor kan det være at deltakerne i studien slått AV AON når dårlig ytelse av modellen var forventet, slik AT AON bare var aktiv for gode forsøk.
dette forslaget er vanskelig å forene, men med nylige rapporter fra vårt laboratorium som viser at å observere både en ekspert og en nybegynnermodell resulterte i bedre læring av et nytt relativt timingsmønster enn å observere enten en nybegynnermodell eller en ekspertmodell alene. Hvis MAN kunne slå AV AON når man ble informert om at en dårlig demonstrasjon vil bli vist( dvs. en nybegynnermodell), ville læring av den blandede observasjonsgruppen ha matchet og ikke overgått ekspertobservasjonsgruppen. Snarere går vi tilbake til vårt første forslag, foreslår vi at EN fw-protokoll hjelper nybegynnere å oppdage og kvantifisere feil i modellens ytelse, noe De vanligvis gjør dårlig (Aglioti et al., 2008; Wright et al., 2010; Ove et al., 2012; Tomeo et al., 2013; Balser et al., 2014; Kristian et al., 2014; Renden et al., 2014). I sin tur kan bedre deteksjon og kvantifisering av modellens ytelse favorisere utviklingen av inverse (Jordan, 1996) og fremovermodeller (Wolpert Og Miall, 1996) av motorstyring.
til slutt er observasjon et kraftig læringsverktøy som er tilgjengelig for alle og krever bare minimalt utstyr som skal brukes. Det er nå godt demonstrert at fordelene med observasjon for å endre den relative timingen (dvs. tempo) av motoriske ferdigheter forbedres når man har tilgang til en rekke forestillinger som spenner fra nybegynnere til eksperter, enten gjennom variable eller blandede observasjonsplaner. Resultatene av denne studien tyder på at disse fordelene er optimalisert hvis observatøren vet på forhånd kvaliteten på ytelsen som hun eller han skal observere under den første observasjonsøkten. Dette kan være svært viktig i en klasseromssammenheng der en lærer / trener vil bruke en videoobservasjonsprotokoll. For eksempel, hvis observatørens intensjon er å lære et bestemt aspekt av en golfsving, er det sannsynlig at resultatet av svingen (dvs. ballflyget) ikke vil bli vist på videoen. Derfor ville observatøren ikke kunne «gjette» modellens ekspertise fra resultatet av svingen, og som vi har vist i denne studien, lære bedre om han eller hun ble informert på forhånd om kvaliteten på det han eller hun skal observere.
Forfatterbidrag
alle forfattere oppført, har gjort betydelige, direkte og intellektuelle bidrag til arbeidet, og godkjent det for publisering.
Finansiering
dette arbeidet ble støttet av Et Discovery Grant (LP) levert Av Natural Sciences And Engineering Research Council Of Canada (stipend nr. 111280-2013).
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
Fotnoter
1. ^For å fastslå at forskjellene i de to retensjonstestene mellom kontrollgruppen og FW-og FB-gruppene resulterte fra en signifikant reduksjon i / CE / av total bevegelsestid, dekomponerte vi i en supplerende analyse gruppemerket faseinteraksjon rapportert i hovedteksten ved å beregne en separat ANOVA for hver gruppe. Resultatene viste at for kontroll-og FB-gruppene var| CE / av total bevegelsestid ikke signifikant forskjellig i henholdsvis fasene. Tvert imot var DET FOR FW-gruppene en signifikant hovedvirkning av fasene, F(2, 86) = 11.60, p < 0,01, np2=0,1, som viste en signifikant reduksjon i| CE / av total bevegelsestid som går fra forprøven til de to retensjonstestene (p < 0,01), som ikke var vesentlig forskjellig fra hverandre (p>0,10).
2. ^Som vi gjorde for| CE / av total bevegelsestid, dekomponerte vi i en supplerende analyse gruppemerket faseinteraksjon rapportert i hovedteksten FOR RMSE av relativ timing ved å beregne en separat ANOVA for hver gruppe. Resultatene viste at FOR kontrollgruppen VAR RMSE av relativ timing ikke signifikant forskjellig over fasene, F(2, 86) = 0,32, p = 0,72, np2=0,01. Tvert imot, for BÅDE FB og FW-gruppene, var det en signifikant hovedeffekt av fasene som viste en signifikant reduksjon I RMSE av relativ timing som gikk fra forprøven til de to retensjonstestene (p < 0,01), som ikke var vesentlig forskjellig fra hverandre (p > 0,10).
3. ^Som vi gjorde i Eksperiment 1, i en supplerende analyse dekomponerte vi gruppemerkingsfaseinteraksjonen rapportert i hovedteksten ved å beregne en separat ANOVA for hver gruppe. Resultatene viste at FOR FB1 – 2-gruppen var| CE / av total bevegelsestid ikke signifikant forskjellig over fasene, F(2, 56) < 1, p = 0,45, np2=0,03. Tvert imot, for BÅDE fw1-2 og FW-FB-gruppene, var det en signifikant hovedeffekt av fasene, som for begge grupper viste en signifikant reduksjon i / CE / av total bevegelsestid som går fra forprøven til de to retensjonstestene (p < 0.01), som ikke skilte seg vesentlig fra hverandre (p> 0,10).
4. ^FOR VE av total bevegelsestid, viste nedbrytingen av gruppemerket faseinteraksjon en signifikant hovedeffekt av faser for alle tre gruppene . For fw1-2-og FB1-2-gruppene viste pos-hoc-sammenligninger en signifikant større VE i pre-test enn i begge retensjonstester (p < 0,01), som ikke skilte seg vesentlig fra hverandre (p>0,30). FOR FW-FB-gruppen VAR VE av total bevegelsestid betydelig større i forprøven enn i 24-h retensjonstesten (p < 0,01).
Abreu, A. M., Macaluso, E., Og Azevedo, R. T. (2012). Handling forventning utover action observation network: en funksjonell magnetisk resonans imaging studie i ekspert basketball spillere. Eur. J. Neurosci. 35, 1646–1654. doi: 10.1111/j.1460-9568. 2012. 08104.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Aglioti, S. M., Cesari, P., Romani, M. ,Og Urgesi, C. (2008). Handling forventning og motor resonans i elite basketball spillere. Nat. Neurosci. 11, 1109–1116. doi: 10.1038 / nn.2182
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Al-Abood, S. A., Davids, K. F. og Bennett, S. J. (2001). Spesifisitet av oppgavebegrensninger og effekter av visuelle demonstrasjoner og verbale instruksjoner i regi elevenes søk under ferdighetstilegnelse. J. Mot. Oppfør deg. 33, 295–305. doi: 10.1080/00222890109601915
PubMed Abstrakt / Fulltekst / Google Scholar
Alaerts, K., Senot, P., Swinnen, S. P., Craighero, L., Wenderoth, N., Og Fadiga, L. (2010a). Kraftkrav for observert objektløfting er kodet av observatørens motorsystem: EN TMS-studie. Eur. J. Neurosci. 31, 1144–1153. doi: 10.1111/j.1460-9568. 2010. 07124.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Alaerts, K., Swinnen, S. P., Og Wenderoth, N. (2010b). Observere hvordan andre løfter lette eller tunge gjenstander: hvilke visuelle signaler formidler kodingen av muskelkraft i primær motor cortex? Neuropsykologi 48, 2082-2090. doi: 10.1016 / j. nevropsykologi.2010.03.029
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Andrieux, M., Og Proteau, L. (2013). Observasjon læring av en motorisk oppgave: hvem og når? Exp. Hjernereses. 229, 125-137. doi: 10.1007 / s00221-013-3598-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Andrieux, M. Og Proteau, L. (2014). Blandet observasjon favoriserer motor læring gjennom bedre estimering av modellens ytelse. Exp. Hjernereses. 232, 3121-3132. doi: 10.1007 / s00221-014-4000-3
PubMed Abstrakt / Fulltekst / Google Scholar
Badets, A., Og Blandin, Y. (2004). Rollen av kunnskap om resultater frekvens i læring gjennom observasjon. J. Mot. Oppfør deg. 36, 62–70. doi: 10.3200 / JMBR.36.1.62-70
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Badets, A., Og Blandin, Y. (2005). Observasjonslæring: effekter av båndbreddekunnskap om resultater. J. Mot. Oppfør deg. 37, 211–216. doi: 10.3200 / JMBR.37.3.211-216
PubMed Abstract | CrossRef Full Text | Google Scholar
Badets, A., Blandin, Y., Wright, D. L., and Shea, C. H. (2006). Error detection processes during observational learning. Res. Q. Exerc. Sport 77, 177–184. doi: 10.1080/02701367.2006.10599352
PubMed Abstract | CrossRef Full Text | Google Scholar
Balser, N., Lorey, B., Pilgramm, S., Naumann, T., Kindermann, S., Stark, R., et al. (2014). Påvirkningen av kompetanse på hjerneaktivering av handlingsobservasjonsnettverket i påvente av tennis og volleyball tjener. Front. Hum. Neurosci. 8:568. doi: 10.3389 / fnhum.2014.00568
PubMed Abstract | CrossRef Full Text | Google Scholar
Bird, G., Og Heyes, C. (2005). Effektoravhengig læring ved observasjon av en fingerbevegelsessekvens. J. Exp. Psychol. Hum. Percept. Utføre. 31, 262–275. doi: 10.1037/0096-1523.31.2.262
PubMed Abstrakt / Fulltekst / Google Scholar
Black, C. B. Og Wright, D. L. (2000). Kan observasjonspraksis legge til rette for feilgjenkjenning og bevegelsesproduksjon? Re. Q. Øvelse. Sport 71, 331-339. doi: 10.1080/02701367.2000.10608916
PubMed Abstrakt / Fulltekst / Google Scholar
Blandin, Y. (1994). Kognitive Prosesser Involvert Under Læring Av Spatio-Temporal Synkronisering Oppgaver Under Ulike Forhold I Praksis Og Observasjon Av En Modell Emne, Upublisert Doktorgradsavhandling. Universitetet I Montreal.
Jørgensen, A., L., L., L., L., L., L., L., L., L., L., L., L. (1999). Kognitive prosesser som ligger til grunn for observasjonell læring av motoriske ferdigheter. Q. J. Exp. Psychol. 52, 957–979. doi: 10.1080/713755856
Full Text | Google Scholar
Blandin, Y., Proteau, L. Og Alain, C. (1994). Om de kognitive prosessene som ligger til grunn for kontekstuell interferens og observasjonell læring. J. Mot. Oppfør deg. 26, 18–26. doi: 10.1080/00222895.1994.9941657
PubMed Abstrakt / Fulltekst / Google Scholar
Jørgen, G., Jørgensen, F., Jørgensen, G., Jørgensen, g., g., m., m., m., m., m., m., m., m., m., m., m., m., m., m., m., m., m., m., et al. (2001). Handlingsobservasjon aktiverer premotoriske og parietale områder på en somatotopisk måte: en fMRI-studie. Eur. J. Neurosci. 13, 400–404. doi: 10.1046/j.1460-9568. 2001. 01385.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Buchanan, J. J., Og Dean, N. J. (2010). Spesifisitet i praksis fordeler læring i nybegynnere modeller og variabilitet i demonstrasjon fordeler observasjonspraksis. Psychol. Res. 74, 313-326. doi: 10.1007 / s00426-009-0254-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Buchanan, J. J., Ryu, Yu, Zihlman, K., Og Wright, D. L. (2008). Observasjonspraksis av relative, men ikke absolutte bevegelsesfunksjoner i en enkelt-lem multi-joint koordinasjonsoppgave. Exp. Brain Res. 191, 157-169. doi: 10.1007 / s00221-008-1512-8
PubMed Abstrakt / Fulltekst / Google Scholar
Candidi, M., Sacheli, L. M., Mega, I., Og Aglioti, S. M. (2014). Somatotopic kartlegging av piano fingering feil i sensorimotor eksperter: TMS studier i pianister og visuelt trent musikalsk naives. Cereb. Cortex 24, 435-443. doi: 10.1093 / cercor / bhs325
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Carroll, W. R., Og Bandura, A. (1990). Representasjonell veiledning av aksjonsproduksjon i observasjonslæring: en årsaksanalyse. J. Mot. Oppfør deg. 22, 85–97. doi: 10.1080/00222895.1990.10735503
PubMed Abstrakt / Fulltekst / Google Scholar
Cisek, P., Og Kalaska, J. F. (2004). Nevrale korrelater av mental øvelse i dorsal premotorisk cortex. Natur 431, 993-996. doi: 10.1038 / nature03005
PubMed Abstrakt | CrossRef Full Tekst | Google Scholar
Cohen, J. (1988). Statistisk Kraftanalyse For Adferdsvitenskapene. Hillsdale, NJ: Lawrence Erlbaum.
Google Scholar
Collier, G. L. og Wright, C. E. (1995). Temporal rescaling av prøve og komplekse rasjoner i rytmisk tapping. J. Exp. Psychol. Hum. Percept. Utføre. 21, 602–627. doi: 10.1037/0096-1523.21.3.602
PubMed Abstrakt / Fulltekst / Google Scholar
E. S., Kraemer, D. J. M., Hamilton, A. F. D., Kelley, W. M., Og Grafton, S. T. (2009). Sensitivitet av action observation network til fysisk og observasjonell læring. Cereb. Cortex 19, 315-326. doi: 10.1093/cercor | bhn083
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Decety, J., Grè, J., Costes, N., Perani, D., Jeannerod, M., Procyk, E., et al. (1997). Hjerneaktivitet under observasjon av handlinger-påvirkning av handlingsinnhold og fagets strategi. Hjerne 120, 1763-1777. doi: 10.1093 / hjerne / 120.10.1763
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Dushanova, J., Og Donoghue, J. (2010). Neuroner i primær motor cortex engasjert under action observasjon. Eur. J. Neurosci. 31, 386–398. doi: 10.1111/j.1460-9568. 2009. 07067.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Frey, S. H. og Gerry, V. E. (2006). Modulering av nevral aktivitet under observasjonell læring av handlinger og deres sekvensielle ordrer. J. Neurosci. 26, 13194–13201. doi: 10.1523 / JNEUROSCI.3914-06.2006
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Gallese V., Fadiga L., Fogassi L., Rizzolatti G. (2002). «Action representation and the inferior parietal lobule,» I Felles Mekanismer I Persepsjon Og Handling: Oppmerksomhet Og Ytelse, Vol. XIX, eds W. Prinz, Og B. Hommel (Oxford: Oxford University Press), 247-266.
Grafton, S. T., Fadiga, L., Arbib, M. A., Og Rizzolatti, G. (1997). Promotor cortex aktivering under observasjon og navngivning av kjente verktøy. Neuroimage 6, 231-236. doi: 10.1006 / nimg.1997.0293
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Hayes, S. J., Elliott, D., Og Bennett, S. J. (2010). Generelle motorrepresentasjoner utvikles under handlingsobservasjon. Exp. Brain Res. 204, 199-206. doi: 10.1007 / s00221-010-2303-6
PubMed Abstrakt / Fulltekst / Google Scholar
Heyes, C. M. og Foster, C. L. (2002). Motor læring ved observasjon: bevis fra en seriell reaksjonstid oppgave. Q. J. Exp. Psychol. 55, 593–607. doi: 10.1080/02724980143000389
PubMed Abstract / CrossRef Full Text / Google Scholar
Hodges, N. J., Chua, R., Og Franks, I. M. (2003). Videoens rolle i å legge til rette for oppfatning og handling av en ny koordinasjonsbevegelse. J. Mot. Oppfør deg. 35, 247–260. doi: 10.1080/00222890309602138
PubMed Abstrakt / Fulltekst / Google Scholar
Hodges, Nj, Williams, A. M., Hayes, Sj, Og Breslin, G. (2007). Hva er modellert under observasjonell læring? J. Sports Sci. 25, 531–545. doi: 10.1080/02640410600946860
PubMed Abstract / CrossRef Full Text / Google Scholar
Jordan, M. I. (1996). «Computational aspects of motor control and motor learning,» i Håndbok For Oppfatning Og Handling: Vol. 2, Motoriske Ferdigheter, eds H. Heuer og S. W. Keele (New York, NY: Academic Press), 71-120.
Kilner, J. M., Neal, A., Weiskopf, N., Friston, K. J., Og Frith, C. D. (2009). Bevis på speilnevroner i menneskelig inferior frontal gyrus. J. Neurosci. 29, 10153–10159. doi: 10.1523 / JNEUROSCI.2668-09.2009
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Lago-Rodrí, A., Cheeran, B., Og Koch, G. (2014). Rollen av speilneuroner i observasjonsmotor læring: en integrativ gjennomgang. Eur. J. Menneskelig Mov. 32, 82–103.
Lee, T. D., Swinnen, S. P. og Serrien, D. J. (1994). Kognitiv innsats og motorisk læring. Quest 46, 328-344. doi: 10.1080/00336297.1994.10484130
Full Text | Google Scholar
Lee, T. D. Og White, M. A. (1990). Påvirkning av en ufaglært modeller praksis tidsplan på observasjons motor læring. Hum. Mov. Sci. 9, 349–367. doi: 10.1016/0167-9457(90)90008-2
Full Text | Google Scholar
Martens, R., Burwitz, L., Og Zuckerman, J. (1976). Modellering effekter på motor-ytelse. Res. Q. 47, 277-291.
PubMed Abstract | Google Scholar
McCullagh, P., Og Caird, J. K. (1990). Korrekt og læringsmodeller og bruk av modellkunnskap om resultater i oppkjøp og oppbevaring av motoriske ferdigheter. J. Menneskelig Mov. Hingst. 18, 107–116.
McCullagh, P. Og Meyer, K. N. (1997). Læring versus riktige modeller: påvirkning av modelltype på læring av en fri-vekt squat lift. Res. Q. Exerc. Sport 68, 56-61. doi: 10.1080/02701367.1997.10608866
PubMed Abstrakt / Fulltekst / Google Scholar
McCullagh, P., Weiss, M. R. og Ross, D. (1989). Modelleringshensyn i motorisk ferdighetstilegnelse og ytelse: en integrert tilnærming. Exerc. Sport Sci. Åp 17, 475-513.
PubMed Abstract | Google Scholar
Oosterhof, N. N., Wiggett, A. J., Diedrichsen, J., Tipper, S. P., Og Downing, P. E. (2010). Overflatebasert informasjon kartlegging avslører tverrmodale visjon-handling representasjoner i human parietal og occipitotemporal cortex. J. Nevrofysiol. 104, 1077–1089. doi: 10.1152 / jn.00326.2010
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Pollock, B. J. Og Lee, T. D. (1992). Effekter av modellens ferdighetsnivå på observasjonsmotor læring. Res. Q. Exerc. Sport 63, 25-29. doi: 10.1080/02701367.1992.10607553
PubMed Abstrakt / Fulltekst / Google Scholar
Renden, P. G., Kerstens, S., Oudejans, R. R. D., Og Cañ-Bruland, R. (2014). Feil eller dykk? Motor bidrag til å bedømme tvetydige foul situasjoner i fotball. Eur. J. Sport Sci. 14 (Suppl. 1), S221-S227. doi: 10.1080/17461391.2012.683813
PubMed Abstrakt / Fulltekst / Google Scholar
Rizzolatti, G., Cattaneo, L., Fabbri-Destro, M., Og Rozzi, S. (2014). Kortikale mekanismer som ligger til grunn for organiseringen av målrettede handlinger og speiler nevronbasert handlingsforståelse. Physiol. Åp 94, 655-706. doi: 10.1152 / physrev.00009.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Rizzolatti, G., Og Fogassi, L. (2014). Speilmekanismen: nylige funn og perspektiver. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 369:20130420. doi: 10.1098 / rstb.2013.0420
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Rohbanfard, H. Og Proteau, L. (2011). Læring gjennom observasjon: en kombinasjon av ekspert-og nybegynnermodeller favoriserer læring. Exp. Brain Res. 215, 183-197. doi: 10.1007 / s00221-011-2882-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Senot, P., D ‘ Ausilio, A., Franca, M., Caselli, L., Craighero, L., Og Fadiga, L. (2011). Effekt av vektrelaterte etiketter på kortikospinal spenning under observasjon av fatte: EN tms-studie. Exp. Hjerne Res. 211, 161-167. doi: 10.1007 / s00221-011-2635-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Ste-Marie, D. M., Law, B., Rymal, A. M., Jenny, O., Hall, C. og McCullagh, P. (2012). Observasjonsintervensjoner for motorisk ferdighetslæring og ytelse: en anvendt modell for bruk av observasjon. Int. Sport Exerc. Psychol. 5, 145–176. doi: 10.1080/1750984x.2012.665076
Full Text | Google Scholar
Tomeo, E., Cesari, P., Aglioti, Sm, Og Urgesi, C. (2013). Lure kickers, men ikke keepere: behavioral og nevrofysiologiske korrelater av falsk handling deteksjon i fotball. Cereb. Cortex 23, 2765-2778. doi: 10.1093/cercor | bhs279
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Vogt, S., Og Thomaschke, R. (2007). Fra visuo-motoriske interaksjoner til imitasjonslæring: atferds-og hjerneavbildningsstudier. J. Sports Sci. 25, 497–517. doi: 10.1080/02640410600946779
PubMed Abstrakt / Fulltekst / Google Scholar
Wolpert, D. M. Og Miall, R. C. (1996). Fremovermodeller for fysiologisk motorstyring. Nevrale Nettverk. 9, 1265–1279. doi: 10.1016 / S0893-6080(96)00035-4
PubMed Abstrakt / Fulltekst / Google Scholar
Wright, D. L., Li, Y. Og Coady, W. (1997). Kognitive prosesser relatert til kontekstuell interferens og observasjonslæring: en replikering Av Blandin, Proteau og Alain (1994). Res. Q. Exerc. Sport 68, 106-109. doi: 10.1080/02701367.1997.10608872
PubMed Abstrakt / Fulltekst / Google Scholar
Wright, M. J., Bishop, D. T., Jackson, R. C., and Abernethy, B. (2010). Functional MRI reveals expert-novice differences during sport-related anticipation. Neuroreport 21, 94–98. doi: 10.1097/WNR.0b013e328333dff2
PubMed Abstract | CrossRef Full Text | Google Scholar