Introduction
ATPは、生物の様々な代謝過程に不可欠な要件です。 生命の継続のためには、ATPの継続的な供給が必要です。 ATPの合成の中断は、有害で生命を脅かす事象を引き起こす可能性があります。
ほとんどの生体系では、ATPは既に存在するADP分子のリン酸化によって作られます。 このリン酸化プロセスは、いくつかの化学エネルギーを必要とする吸熱プロセスです。 このエネルギーは、従属栄養生物で起こるように食物から得られた複雑な化合物を破壊するか、光合成で起こるように光の形で太陽エネルギーを捕獲して利用することによって提供される。
いずれの場合も、ATP合成は化学浸透のプロセスによって起こる。 化学浸透は、イオンの半透膜すなわち浸透を介してそれらの濃度勾配を下にイオンの動きとして定義されます。 この記事では、化学物質理論と、それが細胞のエネルギー通貨であるATPを作るのに役立つメカニズムについて詳細に議論します。
化学物質理論
化学物質理論は1961年にPeter D.Mitchellによって初めて発表されました。 彼は、代謝細胞のATPの大部分は、内部ミトコンドリア膜を横切る電気化学的勾配に蓄積されたエネルギーを利用して合成されることを示唆した。 この電気化学勾配は、高エネルギー分子、NADH、およびFADH2を使用して最初に確立されました。 これらの化合物は、グルコースなどのような食物分子の代謝中に形成された。
酸素代謝の間、それは代謝されてアセチルCoAを形成し、ミトコンドリアマトリックス中でさらに代謝される。 アセチルCoAの分子はクエン酸周期と呼ばれるプロセスの酸化に応じてあります。 このサイクルは、NADやFADのような中間体の還元と結びついています。 還元の結果として形成された高エネルギー中間体(NADHおよびFADH2)は、電子輸送鎖(ETC)に運ばれる。
これらの高エネルギー中間体は、実際には電子のキャリアです。 NADHとFADH2の電子は電子輸送鎖に供与される。 電子が等の下で動くと同時に、内部のmitochondrial膜を渡る電気化学の勾配を作り出すのに使用されている多量のエネルギーは解放されます。 この勾配のエネルギーは、後にプロトンポンプを駆動するために使用され、最終的にADPをATPにリン酸化する。
この化学物質の理論は、当時の科学者の見解に反していたため、すぐには受け入れられませんでした。 電子流のエネルギーは、ATPを作るために直接使用されたいくつかの高エネルギー中間体の形で貯蔵されたと考えられていた。 しかし、時間の経過とともに、科学的証拠は化学物質仮説を証明し始めた。 この理論は受け入れられ、ミッチェルは1978年にノーベル化学賞を受賞した。
ケミオスモティック理論は、ミトコンドリア、葉緑体、および多くの細菌におけるATP合成を説明するようになりました。 これらすべての細胞小器官における化学浸透理論の適用は、この記事の後続の部分で詳細に議論されている。
ミトコンドリアの化学物質
化学物質は、原核生物の細胞呼吸中のATPの主要な供給源である。 このプロセスは生きている細胞のmitochondriaで起こります。 ケミオスモーシスの過程に飛び込む前に、ミトコンドリアの構造を理解してみましょう。
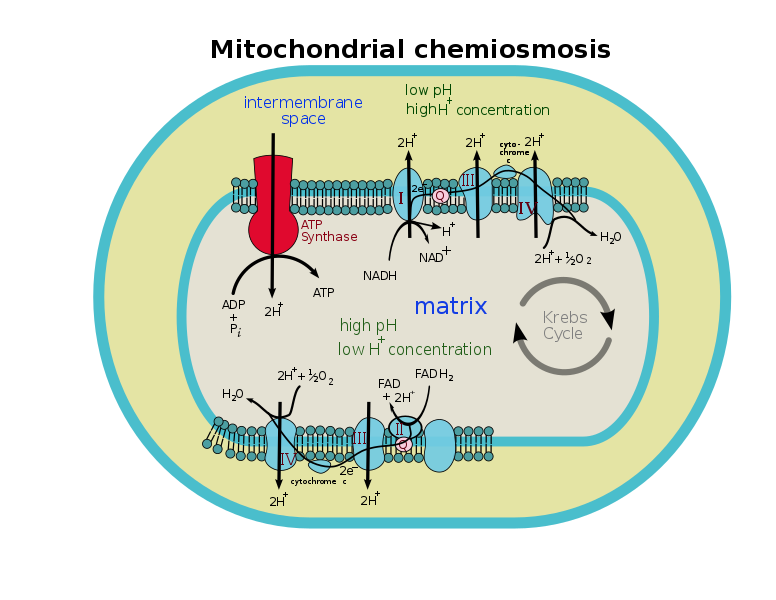
ミトコンドリアの構造
ミトコンドリアは、いくつかの例外を除いて、すべての真核細胞に存在する二重膜結合オルガネラである。 外の膜は内部の膜がさまざまなinfoldingsを示す間、滑らかです。 電子輸送鎖は、ミトコンドリア内膜上に位置する。
電子輸送鎖
電子輸送鎖は、ミトコンドリア内膜に埋め込まれた四つのタンパク質複合体で構成されています。
- 複合体I:NADHデヒドロゲナーゼ、FMN、鉄-硫黄タンパク質から構成されています。
- 複合体II:複合体Iと同じようにコハク酸デヒドロゲナーゼ、FAD、鉄-硫黄タンパク質を有する複合体である。
- 複合体II:シトクロムbとシトクロムc1を有するシトクロム複合体である。 シトクロムは、電子キャリアとして作用するヘムタンパク質である。
- : これは、シトクロムaとシトクロムa3を含む別のシトクロム複合体である。 シトクロムa3は銅含有シトクロムである。 さらに、別の銅含有タンパク質CuAもこの複合体中に存在する。
コエンザイムQは電子輸送鎖のメンバーでもあります。 それは内部のmitochondrial膜で埋め込まれる長いisoprenoid尾を持っているキニーネの派生物です。 それは本質的に遍在しており、ユビキノンとも呼ばれています。 脂質の容解性およびイソプレノイドの構造のために、補酵素Qは内部のmitochondrial膜に沿って自由に動くことができます。 従って、それはまた自由なか移動式電子キャリアとみなされます。
シトクロムcは、膜内空間に存在するシトクロムであり、電子輸送鎖の構成要素でもある。
ATPシンターゼ
電子輸送鎖に加えて、複合体Vと呼ばれる内部ミトコンドリア膜に別の複合体が存在する。 従って、それは別名ATPのシンターゼです。
ATPシンターゼのプロトンチャネルは環と結合している。 陽子がチャネルを通過すると、それらはリングを回転させ、ADPをリン酸化するために使用されるエネルギーが生成される。
膜間空間
それは外側と内側のミトコンドリア膜の間の空間です。 この空間における異なるイオンの濃度は、ミトコンドリアマトリックスとは異なる。 ミトコンドリアマトリックスからのプロトンは、化学浸透のためにこの空間にポンプ輸送され、貯蔵される。
プロセス
ミトコンドリアにおける化学物質プロセスには、以下の手順が含まれます;
- 電子は、NADHやFADH2のような高エネルギー電子キャリアを介して電子輸送鎖に供給されます。 NADHはETCの複合体Iに電子を供給し、FADH2は複合体IIに電子を供給します。
- 電子は電子輸送鎖を下に移動し、かなりの量のエネルギーを解放します。 ETC中の電子の流れは、次の式で表すことができます。:
複合体I->複合体II->コエンザイムQ->複合体III->シトクロムc->複合体IV->酸素
酸素は電子輸送鎖における電子の最終受容体として作用する。
- 水素イオンまたは陽子は、ミトコンドリアマトリックス内ですでに低濃度になっています。 電子によって解放されたエネルギーは、これらの陽子をそれらの濃度勾配に対して膜間空間にポンプ輸送するために使用される。 このようにして、電子のエネルギーは電気化学勾配の形で貯蔵される。
- 陽子が膜間空間に特定の濃度まで集まると、それらはATP合成酵素のプロトンチャネルを通って濃度勾配を下に移動し始める。 このプロセスの間に、彼らはプロトンリングを回転させ、エネルギーを解放する。
- このエネルギーは、ATPシンターゼによって内部ミトコンドリア膜の間質側のATPにADPをリン酸化するために使用されます。
重要性
ミトコンドリアにおける化学物質プロセスは、細胞呼吸を介してエネルギーを得る源である。 このプロセスの障害は、細胞呼吸を介してエネルギーを得ることを不可能にする。
阻害
このプロセスは、電子輸送鎖またはアンカプラタンパク質の任意の阻害剤によって阻害することができる。 ATP合成酵素を通過せずにミトコンドリア間質に入るためのプロトンへの代替パスを提供するアンカプラプロテインチャネル。 電気化学勾配のエネルギーは熱の形で無駄になり、ATPは作られません。 ある薬剤はまたAsprinのようなuncoupler蛋白質として機能します。
葉緑体のケミオスモーシス
葉緑体は光合成独立栄養生物に存在する細胞小器官です。 光励起された電子のエネルギーが暗い反応のためにATPを作るために使用されるとき、オルガネラの化学浸透は光合成の光依存反応の間に起こる。
まず葉緑体の構造を理解しましょう。
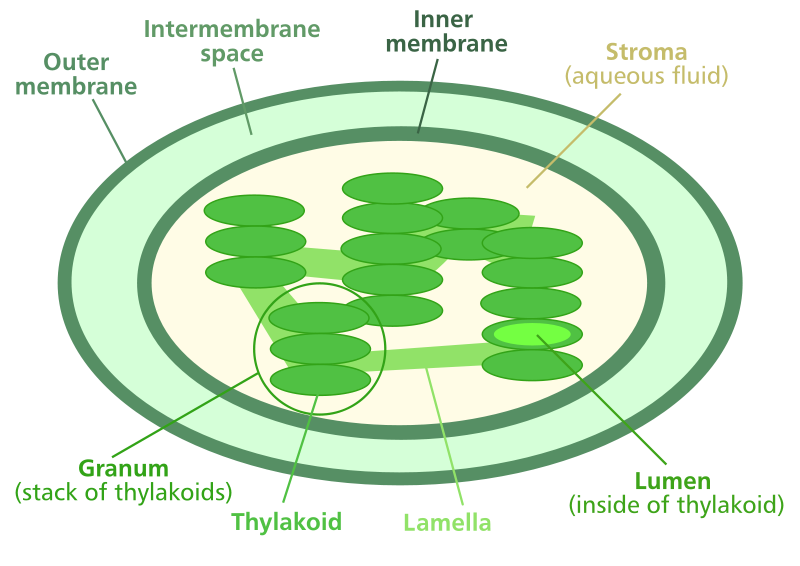
構造
ミトコンドリアと同じように、葉緑体も二重膜細胞小器官です。 しかし、葉緑体の両方の膜は、任意のinfoldingsなしで滑らかです。 葉緑体の間質は細胞小器官の空間の大部分を満たしていた。
チラコイドは、葉緑体の内部に存在するコイン状の構造であり、互いに積み重ねられてグラナを形成する。 チラコイドは、光依存性反応および化学浸透症の部位である。 それらは、チラコイド膜と呼ばれる膜によって結合された内腔で構成されている。
クロロフィル分子と電子輸送鎖の光系はチラコイド膜上に位置している。
電子輸送鎖
チラコイド膜上の電子輸送鎖は、ミトコンドリアに存在するものとは異なる。 それはチラコイド膜上に存在する光システムと結合されている。
光システムは、光エネルギーを集め、クロロフィル分子の電子を励起し、それを電子輸送鎖に移動させるクロロフィル分子のクラスターです。
プラストキノン(Pq)と呼ばれる電子キャリアは、光化学系IIと密接に関連して存在する。
光化学系IIの隣に二つのシトクロムからなるシトクロム複合体が存在している。
次のシリーズは光化学系Iである。プラストシアニン(Pc)と呼ばれる銅含有タンパク質とフェレドキシン(Fd)と呼ばれる鉄含有タンパク質が光化学系Iと密接に接触して存在する。これらのタンパク質は両方とも電子キャリアである。
ATPシンターゼ
光化学系Iの隣にATPシンターゼがある。 これは、内部ミトコンドリア膜に存在するATPシンターゼと同様の構造を有する。 唯一の違いは、プロトンチャネルはチラコイドの内腔に向かって位置し、リン酸化能力を有するF0ドメインは葉緑体の間質に向かって位置するこ
プロセス
チラコイド膜上の化学反応は、光依存反応の間に起こる。 これは、環状電子流と非環状電子流の両方の場合に発生します。
非環状電子流
このプロセスの間、光励起された電子は両方の光系を移動する。 これには、次の手順が含まれます;
- 光の光子が光系に落ちて電子を励起する。
- 光励起された電子は電子輸送鎖を通って移動する。 これらの電子の経路は、次の式で表すことができます:
光化学系I->プラストキノン->シトクロム錯体->プラストシアニン->光化学系II->フェレドキシン->NADP
NADPは電子の最終受容体である。
- 電子が電子輸送鎖を下に移動すると、エネルギーが解放され、葉緑体の間質からチラコイドの内腔に水素イオンを送り込むために使用されます。
- 電子のエネルギーは、チラコイド膜を横切る陽子の電気化学的勾配の形で貯蔵される。
- これらのプロトンは、ATP合成酵素のプロトンチャネルを通過しながら、濃度勾配を下に移動して間質に戻る。 彼らの旅の間、陽子はリングを回転させ、エネルギーを解放する。
- このエネルギーは、葉緑体の間質中のADPをATPにリン酸化するために使用されます。
周期的電子流
周期的な流れでは、光励起された電子は電子輸送鎖を通過し、周期ごとに光化学系Iに戻る。 電子の流れは次のように表されます:
光化学系II->フェレドキシン->シトクロム複合体->プラストシアニン->光化学系II
電子が電子輸送鎖を通過すると、そのエ ATPは、これらの陽子が電子の非周期的な流れのように間質に拡散するときに作られます。
重要性
葉緑体のケミオスモーシスは、光合成の暗い反応のためのATP分子の源です。 化学物質プロセスがATP分子を作ることができない場合、暗い反応は進行することができず、生物はグルコースを製造することができない。 この化学浸透過程は、光合成の過程において主に重要である。 これは、光エネルギーが化学エネルギーに変換され、ATPの分子に高エネルギー結合として保存されるプロセスです。
概要
ケミオスモーシスは、光合成と同様に細胞呼吸におけるATP合成と結合した協奏勾配を下る陽子の動きである。
Peter D.Mitchellは1961年にこの仮説を最初に提案した。 最初は、それは受け入れられませんでした。 しかし、数年後、実験的証拠に基づいて広く受け入れられました。
ケミオスモーシスには、ミトコンドリアと葉緑体に位置する電子輸送鎖が含まれています。
ミトコンドリアにおける化学物質プロセスは、細胞呼吸の間に起こる。
- NADHとFADH2は、ミトコンドリア内膜上のETCに電子を供給する。
- 電子がETCを下に移動すると、陽子は濃度勾配に対して励起されます。
- プロトンはATPシンターゼを通過することによってマトリックスに戻る。
- 陽子はATPを作るために使用されるエネルギーを放出する。
葉緑体の化学物質プロセスは、光合成の間に起こります。
- 光励起された電子はチラコイド膜上のETCを下に移動する。
- 電子のエネルギーは、間質からチラコイド内腔に陽子を送り込むために使用されます。
- プロトンが間質に戻ると、ATPシンターゼを通過する。
- エネルギーまたは陽子は、ATPシンターゼによってATPを作るために使用されます。
このプロセスは、光依存反応における電子の周期的および非周期的な流れの両方の間に起こる。