acumularea de dovezi sugerează că întreruperile semnalizării fosforilării/defosforilării sunt asociate cu comportamentele patologice ale celulelor. Proiectele fosfo-proteome au identificat aproape 300.000 de reziduuri Ser/Thr/Tyr fosforilate în proteinele celulare . În paralel, Proiectul genomului uman a descoperit aproximativ 500 de proteine kinaze. Cu toate acestea, un număr limitat de protein kinaze au fost atribuite fiecărui situs de fosforilare.
testul de fosforilare în gel a fost o metodă convenabilă pentru detectarea activităților protein kinazei în probele solubilizate cu SDS. Această metodă a fost aplicată pentru identificarea CaMKII, MYPT1-K (sau ZIPK) și altele . În acest protocol de testare, protein kinazele sunt încărcate pe un gel SDS-poliacrilamidă încorporat cu o polipeptidă de substrat și apoi supuse renaturării, fosforilării folosind ATP radioactiv, urmată de autoradiografie pentru a detecta substratul fosforilat. În plus, autofosforilarea kinazelor a fost detectată pe membrana de nitroceluloză sau fluorură de poliviniliden (PVDF) utilizând ATP radioactiv . Materialul radioactiv poate fi o limitare pentru utilizatori, din cauza reglementărilor de siguranță. În plus, autoradiogramele proteinelor fosforilate nu au informații despre locurile de fosforilare.
aici, am optimizat un test alternativ non-radioactiv la testul tradițional de fosforilare în gel. Numit activitate kinază-etichetat Western blotting (KAT-WB), aceasta detectează cu succes Tyr-kinazele care se autofosforilează, precum și Ser/Thr-kinazele care fosforilează o proteină substrat de interes la un anumit loc folosind anticorpi specifici fosfo (P).
Figura 1 prezintă datele KAT-WB pentru testul de autofosforilare a Tyr-kinazei. Pentru a face bloturi duplicate, două alicote dintr-un extract au fost încărcate pe un gel de poliacrilamidă SDS, supus electroforezei, iar apoi proteinele au fost transferate pe o foaie de membrană PVDF (0,2 centimm). Membrana a fost tratată secvențial cu 2-propanol tamponat, 6,0 m clorhidrat de guanidină (Gu-HCl), 3,0 M gu-HCl, 0,1 m gu-HCl și tampon de renaturare (vezi Protocolul suplimentar). După o etapă de renaturare peste noapte la 4 inkt C, membrana a fost tăiată longitudinal pentru a genera două piese duplicate (Figura 1). O bucată de membrană a fost incubată timp de 60 min cu MgATP de 1 mM. Cealaltă bucată de membrană a fost incubată fără MgATP, ca un control. Ambele foi au fost supuse simultan imunodetecției cu anticorp anti-P-Tyr (clonă PY20 la diluție 1:5000). După cum se arată în Figura 1, mai multe benzi P-Tyr au fost detectate la 100, 70, 45 și 35 kDa în membrana de control (stânga sus). Aceste proteine P-Tyr au fost probabil deja fosforilate înainte de extracție. Incubația cu MgATP după separare a crescut fosforilarea Tyr a unei proteine la 45 kDa în lizatele musculare cardiace (Figura 1, dreapta sus, banda 2), indicând autofosforilarea reziduurilor Tyr în această kinază pe membrană după renaturare. Figura 1 prezintă densitogramele benzii 2 (-/+ MgATP). Pentru a verifica încărcarea echivalentă a proteinelor pe cele două pete, filtrele au fost supuse colorării Coomassie (Figura 1, partea de jos). O notă tehnică este că, în cursul dezvoltării metodei, am constatat că tratamentele cu 2-propanol tamponat și 6m gu-HCl au fost necesare pentru detectarea activităților kinazei. Prin urmare, KAT-WB a detectat o kinază Tyr de 45-kDa.

proteinele totale ale musculaturii scheletice de șoarece (31%), mușchiului cardiac (28%) și celulelor Neuro2a (27%) au fost supuse la KAT-western blotting. După renaturare, membrana a fost tăiată longitudinal în două și fiecare parte a fost incubată timp de 60 min la 37 centimetric C în absența (stânga) sau prezența (dreapta) a 1 mm ATP. Fosforilarea Tyr a fost detectată printr-o metodă electrochimiluminiscentă convențională folosind anticorp anti-P-Tyr (top). După imunoblotting, proteinele totale de pe fiecare pată au fost vizualizate prin colorarea Coomassie (partea de jos). Numerele din dreapta indică greutățile moleculare ale proteinelor marker (în kDa). O casetă din dreapta indică densitograme ale benzii 2 în panourile din stânga și din dreapta. Nivelurile de luminozitate și contrast ale bloturilor sunt identice.
o altă versiune a KAT-WB utilizează fosforilarea specifică site-ului a unei proteine de substrat absorbite pe membrana PVDF în apropierea kinazei. Am folosit inhibitorul holoenzimei fosfatazei-1 (PHI-1) ca proteină de interes, a cărei fosforilare la Thr57 transformă această proteină celulară într-un inhibitor al fosfatazei Ser/Thr de tip 1 . Figura 2a prezintă datele KAT-WB utilizând ca martor pozitiv ZIPK recombinant, o kinază Ser/Thr cunoscută pentru fosforilarea PHI-1 recombinant la Thr57 . După denaturarea și procesul de renaturare în trepte, membrana a fost îmbibată peste noapte la 4 inkt C în tamponul de renaturare suplimentat cu PHI-1 recombinant de tip sălbatic (WT) sau o versiune nefosforilabilă (T57H) la 0,2 mg/ml, iar apoi reacția de fosforilare a fost rulată cu MgATP. PHI-1 fosforilat pe membrană a fost detectat folosind anticorp anti-P-PHI-1 (Thr57), iar acest lucru a relevat o bandă a zipk recombinant. Cu toate acestea, doar un semnal slab a fost detectat pe membranele de control fără PHI-1 sau cu versiunea T57H a PHI-1. Aceste date sugerează că kinazele renaturate de pe membrană sunt capabile să fosforileze substraturi care sunt, de asemenea, absorbite pe membrană. KAT-WB a fost aplicat pentru a detecta mai multe kinaze PHI-1 în țesuturi și celule (figura 2b). Extractele din scheletul șobolanului, mușchii cardiaci și celulele Neuro2a au fost supuse testului KAT-WB. Comparativ cu controlul no-ATP (stânga, sus), semnalele P-PHI-1 au apărut pe blotul incubat cu 1 mM MgATP (dreapta, sus). Benzi proeminente de P-PHI-1 au fost detectate în lizatele celulelor Neuro2a la 100, 60 și 49 kDa (banda 3, vârful săgeții deschise). Cu toate acestea, nu au fost evidente benzi pentru P-PHI-1 pe banda cu extracte de mușchi scheletici și cardiaci (benzile 1 și 2). Semnalul P-PHI-1 nu a fost îmbunătățit prin etapa de fosforilare fără PHI-1 (Figura 2, fund, gol: BLK). Aceste rezultate arată că există mai multe kinaze PHI-1 exprimate în celulele neuronale, dar nu în mușchii scheletici sau cardiaci. Acest lucru ne oferă un instrument puternic pentru detectarea mai multor kinaze specifice substratului în celule și țesuturi. O notă tehnică este că renaturarea peste noapte cu PHI-1 a fost necesară pentru detectarea semnalelor de fosforilare și că o concentrație mai mare de PHI-1 nu a crescut intensitatea colorării (nu a fost prezentată).
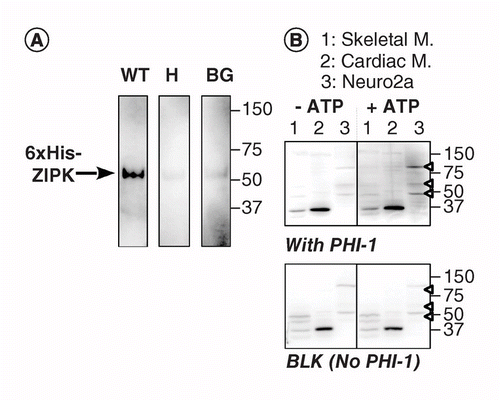
(a) bloturile Triplicate au fost preparate folosind zipk recombinant parțial purificat din lizați de celule care exprimă o versiune etichetată his cu 6%. După electroblotting și procesul de denaturare–renaturare, membrana a fost incubată peste noapte cu proteine recombinante PHI-1 (WT: stânga; T57H: centru) la 0,2 mg/ml sau fără PHI-1 (BG). Fiecare membrană a fost apoi incubată cu 1 mm ATP la 37 C și supusă imunodetecției folosind anticorp anti-P-PHI-1 (T57). (B) au fost preparate două bloturi duplicate (de sus și de jos) utilizând probe totale de proteine, conform descrierii din Figura 1. PHI-1 Recombinant (0,2 mg/ml) a fost adăugat la tamponul de renaturare pentru o pată (top). BLK indică pata fără tratamentul recombinant PHI-1. După incubare în absența (stânga) și prezența (dreapta) a 1 mm ATP, bloturile au fost supuse imunodetecției folosind anti-P-PHI-1 (Thr57). Săgețile din dreapta indică locațiile P-PHI-1 în panoul din dreapta sus. Nivelurile de luminozitate și contrast ale bloturilor sunt identice.
BG: fundal; WT: tip sălbatic.
pe baza acestor date, propunem ca KAT-WB să fie capabil să detecteze autofosforilarea kinazelor Tyr și fosforilarea specifică site-ului proteinelor substratului selectiv de interes prin mai multe protein kinaze prezente în țesuturi și celule. KAT-WB împărtășește în esență avantajele testului tradițional de fosforilare în gel, care detectează activitățile kinazei într-un amestec brut, cum ar fi extractele celulare, expunând dimensiunile lor moleculare . Un alt avantaj al acestor două teste este că modificările post-translaționale (PTM) ale kinazelor din celule pot fi păstrate la un moment dorit în condiții specifice, deoarece probele pentru aceste teste sunt denaturate rapid cu tamponul, inclusiv SDS, eliminând fosforilarea și defosforilarea în extracte . Majoritatea protein kinazelor sunt reglate prin PTM. KAT-WB este capabil să monitorizeze fluctuațiile activităților kinazei, chiar dacă PTM-urile de reglementare nu au fost încă caracterizate. KAT-WB nu este în măsură să detecteze fiecare kinază din extractele celulare, deoarece numai enzimele monomerice care renaturează eficient în condițiile utilizate prezintă activitate. Cu toate acestea, modificările KAT-WB ar trebui să poată detecta kinazele reglate de calmodulină și cicline, așa cum s-a făcut prin testul de fosforilare în gel . Îmbunătățirea eficienței renaturării și o combinație cu imunoprecipitarea/imunodepletarea cu anticorpi pentru kinaze pot extinde utilitatea abordării.
este de remarcat faptul că, pe lângă fosforilare, principiul protocolului KAT-WB poate fi aplicat pentru detectarea altor enzime PTM, cum ar fi acetilazele, metilazele și ubiquitin ligazele, atunci când sunt disponibili anticorpi specifici PTM. Odată cu dezvoltarea anticorpilor care recunosc PTM-urile specifice site-ului, obiectivele de cercetare acoperite de western blotting etichetate cu activitate vor fi extinse.
contribuții ale autorului
M Eto, s Katsuki, y Tanaka și K Takeya au conceput și proiectat experimentele. S Katsuki, y Tanaka și K Takeya au efectuat testele. M Eto, s Katsuki, y Tanaka și K Takeya au efectuat analiza datelor. M Eto, s Katsuki, y Tanaka și K Takeya au scris lucrarea.
mulțumiri
autorii ar dori să-i mulțumească dlui Brautigan pentru corectarea manuscrisului și pentru sugestii valoroase.
financiar & concurente interese divulgarea
această lucrare este susținută de JSPS KAKENHI (JP19K06573 și JP19H03125) și Societatea pentru cercetare pe gust Umami, fonduri de la Imabari city și prefectura Ehime, și un cadou de la H Awagi. Autorii nu au nicio altă afiliere sau implicare financiară relevantă cu nicio organizație sau entitate care are un interes financiar sau un conflict financiar cu subiectul sau materialele discutate în manuscris, în afară de cele dezvăluite.
nu a fost utilizată nicio asistență scrisă în producerea acestui manuscris.
Acces Deschis
această lucrare este licențiată sub Licența Atribuire-Necomercial-NoDerivatives 4.0 Unported. Pentru a vizualiza o copie a acestei licențe, vizitați http://creativecommons.org/licenses/by-nc-nd/4.0/
- 1. Hornbeck PV, Zhang B, Murray B și colab. PhosphoSitePlus, 2014: mutații, PTM și recalibrări. Acizi Nucleici Res. 43, D512-D520 (2015).Crossref, Medline, CAS, Google Scholar
- 2. Celenza JL, Carlson M. Renaturarea activității protein kinazei pe bloturile proteice. Metode Enzimol. Academic Press, Cambridge Massachusetts, 34, 423-430 (1991).Crossref, Google Scholar
- 3. Hutchcroft JE, Anostario M, Harrison ML et al. Renaturation and assay of protein kinases after electrophoresis in sodium dodecyl sulfate–polyacrylamide gels. Methods Enzymol. Academic Press,Cambridge Massachusetts, 33, 417–423 (1991).Crossref, Google Scholar
- 4. Kameshita I, Fujisawa H. A sensitive method for detection of calmodulin-dependent protein kinase II activity in sodium dodecyl sulfate-polyacrylamide gel. Anal. Biochem. 183(1), 139–143 (1989).Crossref, Medline, CAS, Google Scholar
- 5. Shackelford DA, Zivin JA. Renaturarea activității protein kinazei dependente de calciu/calmodulină după transferul electroforetic din gelurile de sodiu dodecil sulfat-poliacrilamidă în membrane. Anal. Biochem. 211(1), 131–138 (1993).Crossref, Medline, CAS, Google Scholar
- 6. Macdonald JA, Borman MA, Muranyi A și colab. Identificarea kinazei asociate miozinei fosfatazei musculare netede endogene. Proc. Natl Acad. Sci. SUA 98(5), 2419-2424 (2001).Crossref, Medline, CAS,Google Scholar
- 7. Wooten MW. Testul kinazei în gel ca metodă de identificare a substraturilor kinazei. Sci. STKE 2002 (153), pl15 (2002).Medline, Google Scholar
- 8. Celenza J, Carlson M. o genă de drojdie care este esențială pentru eliberarea din reprimarea glucozei codifică o protein kinază. Știință 233(4769), 1175-1180 (1986).Crossref, Medline, CAS, Google Scholar
- 9. Eto M, Karginov A, Brautigan dl. Un nou inhibitor al fosfoproteinelor de tip proteic-1 fosfatază holoenzime. Biochimie 38(51), 16952-16957 (1999).Crossref, Medline, CAS, Google Scholar
- 10. Macdonald JA, Eto M, Borman MA și colab. Fosforilarea dublă Ser și Thr a CPI-17, un inhibitor al miozinei fosfatazei, de către kinaza asociată MYPT. FEBS Lett. 493(2–3), 91–94 (2001).Crossref, Medline, CAS, Google Scholar
- 11. Eto M, Leach C, Tountas NA și colab. Inhibitori ai fosfoproteinei proteinei fosfatazei-1. Metode Enzimol. 366, 243–260 (2003).Medline, CAS, Google Scholar
- 12. Rousell J, Haddad El B, Lindsay MA și colab. Reglarea expresiei genei receptorului muscarinic m2 prin factorul de creștere derivat din trombocite: implicarea protein kinazelor reglate de semnal extracelular în procesul de reglare în jos. Mol. Pharmacol. 52(6), 966–973 (1997).Crossref, Medline, CAS, Google Scholar