- genomi ai tulpinilor de Finegoldia
- împărțirea tulpinilor Finegoldia în două clade distincte pe baza comparației genomului
- eterogenitatea specifică tulpinii Finegoldia sp
- factori de interacțiune a gazdei și factori de virulență (presupuși)
- genomurile Finegoldia codifică factorii CAMP care sunt funcționali
- două sisteme de teste biochimice nu pot diferenția fără echivoc F. magna de „F. nericia”
- microscopia relevă diferențe de suprafață între F. magna și „F. nericia”
genomi ai tulpinilor de Finegoldia
în total 17 genomi ai Finegoldia sp. au fost analizate și comparate. Zece tulpini au fost recent secvențiate de grupul nostru15; au fost izolate de la pacienți cu infecții ortopedice asociate implantului articular în Suedia. Șapte genomi au fost anterior secvențiate de alții, inclusiv genomul închis al tulpinii de tip ATCC 2932814. Caracteristicile genomului tuturor tulpinilor de Finegoldia secvențiate până acum sunt rezumate în tabelul 1. Adnotarea folosind Prokka a detectat între 1570 și 1906 secvențe de codificare (CDS) pe genom, cu o medie de 1760 CDS pe tulpină.
împărțirea tulpinilor Finegoldia în două clade distincte pe baza comparației genomului
am observat anterior o eterogenitate mare în rândul genomilor izolaților Finegoldia15. S-a făcut o aliniere nucleu-genom a tuturor celor 17 genomi și s-au numit polimorfisme nucleotidice unice (SNP) în genomul de bază. În total, 126.647 SNP-uri cu genom de bază au fost identificate și utilizate pentru reconstrucția filogenomică, dezvăluind că cei 17 genomi pot fi grupați în două clade distincte (Fig. 1). O analiză suplimentară care determină identificarea nucleotidelor medii (ANI) a genomului de bază a arătat că cele două clade prezintă un ANI de 90,7%, cu o variație ANI specifică tulpinii de la 90,2% la 91,2%. Un astfel de ANI scăzut indică faptul că cele două clade reprezintă specii individuale16. O cladă conține cele mai multe tulpini secvențiate și studiate anterior, inclusiv ATCC29328 și ALB8; poate fi considerată clada clasică F. magna, cu un ani intraclade de 96,6% (variație 95,1–97,4%). Cealaltă cladă cu un ani intraclade de 94,8% (93,4–96.6%) conține o altă tulpină ATCC (ATCC 53516) și în principal tulpini izolate de la pacienții de la Spitalul Universitar Ectrebro, Suedia15. În continuare, tulpinile acestei clade sunt denumite în mod provizoriu „Finegoldia nericia” (Latină pentru N Eckrke, Regiunea Suedeză, unde au fost izolate majoritatea tulpinilor acestei specii).

Arborele Filogenomic bazat pe SNP-urile genomului de bază ale tuturor celor 17 genomi secvențiați ai tulpinilor Finegoldia. Programul Parsnp a fost folosit pentru a alinia genomul de bază și pentru a apela SNP-urile. Genomul de bază este reprezentat de 43% din genomul de referință (ATCC29328); un total de 126.647 SNP-uri fiabile ale genomului de bază au fost utilizate pentru a reconstrui o filogenie a genomului întreg. Genomurile pot fi atribuite la două clade principale; o cladă reprezintă tulpinile F. magna, iar cealaltă cladă constă din tulpini ale unei specii noi, numită aici provizoriu „F. nericia”.
o comparație a secvențelor ARNr 16S ale tuturor izolatelor a relevat un grad ridicat de similitudine generală (>99%), dar a detectat o separare a izolatelor F. magna de izolatele „F. nericia”, datorită 5 la 9 „F. nericia ” – SNP-uri specifice (figura S1). De asemenea, a relevat o diversificare mai mare între izolatele „F. nericia” comparativ cu F. magna. În continuare, am analizat secvențe suplimentare de ARNr 16S aparținând Finegoldia sp. stocate în GenBank (figura S2). Acest lucru a arătat că majoritatea secvențelor stocate se grupează cu „F. nericia”, în special un grup de Finegoldia sp. tulpini izolate din biofilme de stent biliar17. Din nou, este detectată o diversificare mai mare între izolatele „F. nericia”. În continuare, am analizat datele microbiomului uman stocate în baza de date IMG (Integrated microbian Genomes and microbiome samples) pentru prezența secvențelor F. magna și „F. nericia”. 3287 și 7068 secvențe de codificare ale F. magna ATCC29328 și, respectiv, „F. nericia” ATCC 53516, au putut fi identificate în cele 875 de seturi de date microbiome umane asamblate prezente în IMG (datele nu sunt prezentate); cele mai mari apariții de Finegoldia au fost detectate la specimenele de Cute retroauriculare. Luate împreună, aceste analize sugerează că printre izolatele Finegoldia asociate omului „F. nericia” pare a fi mai răspândită decât F. magna.
eterogenitatea specifică tulpinii Finegoldia sp
pentru a clarifica structura populației genului Finegoldia, au fost efectuate analize comparative a 12 genomi (toate cele zece tulpini nou secvențiate și cele două tulpini ATCC), incluzând patru tulpini ale cladei F. magna și opt tulpini ale cladei „F. nericia”. O comparație vizuală a celor 12 genomi a evidențiat diferența de identitate nucleotidică dintre cele două clade (Fig. 2). Interesant este că plasmida Găsită în ATCC 29328 pare a fi specifică pentru această tulpină, deoarece nicio altă tulpină secvențiată nu conține o secvență similară (Fig. 2A).

Compararea genomului a 12 tulpini aparținând genului Finegoldia. Cele două inele cele mai interioare reprezintă conținutul G + C (negru) și GC-oblic (violet/Verde). (A) tulpina de referință este ATCC29328 (F. magna); rețineți identitatea nucleotidică ridicată (vizualizată de culoarea roșu închis) a celor trei genomi interiori (aparținând tuturor lui F. magna), iar cea inferioară se identifică la opt tulpini aparținând „F. nericia” (culoarea roșu pal a celor opt inele exterioare); de la interior la exterior: tulpinile 07T609, 08t492, 09t408, 09t494, 12t272, 12t273, 12t306, CCUG 54800, T151023, T160124 și ATCC 53516. Regiunea mare specifică ATCC29328 din regiunea din stânga sus reprezintă plasmida pFMC. (B) tulpina de referință este 09T494 („F. nericia”). Cele șapte genomi” F. nericia ” (inele interioare) au o nucleotidă mai mare identifica (reprezentată de culoarea roșie mai închisă), comparativ cu cele patru genomi F. magna (inele exterioare); din interior spre exterior: tulpinile 12T272, ATCC 53516, 12t306, 12t273, CCUG 54800, T151023, T160124, ATCC29328, 07t609, 08t492 și 09T408. Nu există semne de regiuni genomice specifice „F. nericia”, dar rețineți cele cinci grupuri mari specifice tulpinii.
nu au fost detectate diferențe semnificative în dimensiunea genomului și numărul de CDS între cele două clade. În plus, nu au fost detectate regiuni genomice specifice cladei (adică F. magna sau „F. nericia”), așa cum s-a judecat din analiza BRIG (Fig. 2) și, de asemenea, confirmată de o abordare blastică bidirecțională a tuturor CDS folosind ProteinOrtho (tabelul S1). Această abordare a detectat 1202 ortologi împărțiți de toți cei 12 genomi, adică în medie 68% Din CDS-urile fiecărei tulpini fac parte din proteomul de bază. Se găsesc doar foarte puține gene specifice cladei. În schimb, pot fi găsite multe regiuni împărtășite doar de câteva tulpini, de exemplu pentru tulpina 09T494 (vezi și Fig. 2B). În plus, sunt prezente regiuni specifice tulpinii care codifică în total 1016 CDS specifice tulpinii, cu o gamă largă între tulpini, de exemplu 187 și 22 CDS specifice tulpinii sunt detectate în tulpina ATCC29328 și, respectiv, T151023 (tabelul S1). Printre funcțiile specifice tulpinii se numără diverse funcții de transport (Fier, pompe de eflux, transportori ABC oligopeptidici), proteine de suprafață, Sisteme de modificare a restricțiilor, funcții de transfer conjugativ, sinteza polichetidelor, bacteriocine, funcții legate de fag, determinanți ai rezistenței la antibiotice și sisteme CRISPR/cas.
interesant, diferențele în sistemele CRISPR/cas au fost detectate între tulpinile de F. magna și „F. nericia” (tabelul s2). Șapte din nouă F. tulpinile magna conțineau una până la patru matrice CRISPR pe tulpină, conținând în medie 12 distanțiere (variind de la 3 la 38 distanțiere pe tulpină). Repetarea „GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC” a fost specifică tulpinilor de F. magna. În schimb, tulpinile „F. nericia” purtau o singură matrice CRISPR pe tulpină care conținea în medie 53 de distanțiere (interval de la 14 la 87 pe tulpină). Fiecare tulpină conținea un set individual de distanțieri în matricea lor CRISPR, eterogenitatea specifică tulpinii și indicând faptul că fiecare tulpină are o istorie evolutivă unică.
pentru a evalua dacă astfel de funcții specifice tulpinii au fost dobândite pe orizontală, am analizat semnele mobilității/achiziției lor folosind programul IslandViewer (figura S3). Acest program a prezis 141 (tulpina 09T408) la 408 (tulpina 12t273) gene situate în insule genomice care sunt susceptibile de a fi dobândite pe orizontală, inclusiv multe dintre genele specifice tulpinii menționate mai sus implicate în specializare, fitness, supraviețuire și competiție interspecifică (tabelul S3).
factori de interacțiune a gazdei și factori de virulență (presupuși)
mai mulți factori de interacțiune a gazdei au fost descriși pentru F. magna 4,5. Au fost identificați pili dependenți de sortază care sunt importanți pentru adeziune și colonizare13. Un locus genomic care codifică genele pentru subunitatea pilus Fmp1, subunitatea putativă Fmp2 și trei sortaze este conservat între toate genomii secvențiați (Fig. 3A). Există variații specifice tulpinii în sus și în aval de acest locus, adică. unele tulpini posedă gene suplimentare în amonte de fmp1 și în aval de locusul sortazei, inclusiv o genă care codifică o proteină din familia hemolizinei III. Interesant este că există diferențe specifice tulpinii în ceea ce privește fmp1. Aproape fiecare tulpină poartă o variantă individuală și o analiză filogenetică a Fmp1 la nivelul proteinei nu a arătat nicio corelație cu analiza filogenomică a genomului de bază (Fig. 3B). În schimb, Fmp2 este foarte conservat în rândul tuturor tulpinilor. O altă proteină, codificată în amonte de fmp1, adăpostește domenii de legare a colagenului și fibrinogenului; domeniul de legare a colagenului (COG4932) este prezent și în Fmp2. Propunem ca această proteină să aparțină și locusului pilus și să denumească gena corespunzătoare fmp3.

loci genomici care codifică Pili dependenți de Sortază în genomii tulpinilor Finegoldia. (A) organizarea genomică a locilor în tulpinile F. magna (roșu) și „F. nericia” (Albastru deschis). Cele trei gene sortaze sunt reprezentate în verde deschis (etichetate „2”); gena pentru subunitatea pilus majoră Fmp1 este prezentată în roșu (etichetată „1”) și fmp2 este descrisă în portocaliu (etichetată „3”). În amonte de fmp1 și în aval de genele sortazei în unele tulpini sunt prezente gene suplimentare (etichetate „8”, proteina din familia hemolizinei III și „10”, proteina ipotetică). O proteină presupusă de legare a colagenului, Fmp3 (etichetată „4”), este codificată în amonte de fmp1. (B) arborele filogenetic al fmp1 al tulpinilor Finegoldia. Majoritatea tulpinilor posedă o variantă individuală Fmp1. Istoria evolutivă a fost dedusă folosind metoda probabilității maxime, realizată în MEGA7.
proteina L este probabil cel mai bine studiat factor al F. magna 8. Este un superantigen care vizează celulele B și imunoglobulinele. A fost detectat în aproximativ 10% din izolatele Finegoldia testate până acum, dar variază ca mărime și număr de fragmente de legare a antigenului (Fab)-domenii de legare (domeniul B1, pfam02246)4,8,18. O căutare blastică cu proteina l studiată din tulpina 312 (locus ID: M86697, 719 aa) relevă câteva proteine foarte similare (identitate proteică peste 50%) cu dimensiuni diferite (603 până la 1417 aa) în câteva tulpini, adică. tulpinile CCUG54800, T161024, ATCC53516, 12T273 și 12t306 (Fig. 4A). Toate acestea sunt tulpini „F. nericia”, ceea ce indică faptul că superantigenul cu celule B este mai răspândit la această specie. Factorul asemănător proteinei l excepțional de mare (1417 aa) din tulpina 12t306 posedă pe lângă două domenii B1 și două module GA care sunt cunoscute din proteinele care leagă albumina peptostreptococică (vezi mai jos). Multe proteine suplimentare cu similitudine scăzută cu proteina L, dar fără a conține domenii B1 aparente sunt codificate în tulpinile Finegoldia secvențiate de genom (tabelul S4A); din nou, există variații specifice tulpinii și câteva tulpini, adică 09T408, 09t494 și T151023 nu posedă astfel de omologi.

loci genomici care codifică proteinele l și omologii FAF în tulpinile Finegoldia. (A) căutările blastice cu proteina l caracterizată din tulpina 312 (locus ID: M86697) au fost efectuate în tulpini Finegoldia secvențiate de genom și sunt prezentați omologi (în roșu, etichetați „1”) cu similitudine ridicată (>50% identitate proteică) (a se vedea, de asemenea, tabelul S4A). Numai tulpinile „F. nericia”, dar nu F. tulpinile magna conțin astfel de omologi cu lungimi diferite. (B) căutările de Blast cu proteina FAF studiată a tulpinii ALB8 au găsit omologi de lungime întreagă (în roșu, etichetați „1”) în trei tulpini F. magna (roșu) și patru tulpini „F. nericia” (Albastru deschis). Gena FAF este încadrată în cele două tulpini” F. nericia ” T151023 și 12t272.
s-a constatat că Finegoldia magna se leagă de albumina umană9. O proteină peptostreptococică care leagă albumina (PAB) a fost studiată la tulpina ALB8; activitatea sa este legată de o rată de creștere accelerată9. Proteina PAB din tulpina ALB8 (locus ID: X77864) conține module GA, domeniul de legare a albuminei; structura modulului GA a fost rezolvată19. O căutare explozivă a arătat că un omolog PAB apropiat nu poate fi găsit în alte genomuri finegoldia secvențiate (tabelul S4B). În schimb, există mai mulți omologi care împărtășesc până la 50% identitate proteică; asemănările omologilor din tulpinile F. magna sunt în medie mai mari comparativ cu omologii din tulpinile „F. nericia”. În ceea ce privește proteinele care conțin modulul GA, în total 16 proteine sunt codificate în cele 12 genomi Finegoldia, indicând o anumită redundanță a proteinelor care leagă albumina (tabelul S4C). Majoritatea tulpinilor codifică două proteine cu module GA, cu excepția tulpinii 09T408 (F. magna) și T151023 („F. nericia”).
s-a demonstrat că proteaza sufa subtilizină atașată la peretele celular degradează mai multe proteine umane, cum ar fi colagenul IV, fibrinogenul și peptidele antimicrobiene,cum ar fi LL-37 și MIG/CXCL910,20, 21. Căutările Blast au stabilit că majoritatea tulpinilor Finegoldia poartă un omolog SufA care este foarte similar (identitate proteică > 75%) cu SufA studiat din tulpina ALB8 (locus ID: DQ679960). Excepțiile sunt tulpinile 07T609, 09T408 și CCUG54800. Un al doilea omolog SufA este prezent în majoritatea tulpinilor; are o identitate proteică medie de 26% (tabelul S4D).
un alt factor important de interacțiune a gazdei este FAF (factorul de adeziune F. magna). Aproximativ 90% din tulpinile F. magna produc acest factor, care este responsabil pentru aglomerarea bacteriilor și mediază legarea la membrana bazală prin legarea la BM-4011. Am constatat că șapte (trei tulpini F. magna și patru tulpini „F. nericia”) din cele 12 tulpini codifică un omolog FAF similar cu cel studiat la tulpina ALB8 (Fig. 4B). Există diferențe specifice tulpinii și mutațiile frameshift sunt observate la tulpinile” F. nericia ” T151023 și 12t272. Nici un omolog nu poate fi găsit în tulpinile 09T408, 09t494 și 12t306 (tabelul S4E).
genomurile Finegoldia codifică factorii CAMP care sunt funcționali
am căutat în genom alți factori presupuși de interacțiune a gazdei și am găsit gene care codifică factorii Christie-Atkins-Munch-Petersen (CAMP). Factorii CAMP pot acționa ca co-hemolizine și pot explica așa-numita reacție CAMP, Liza sinergică a eritrocitelor ovine de către Staphylococcus aureus sfingomielinaza C (beta-toxină) și un factor CAMP22.
exploatarea genomului a identificat două gene care codifică factorii CAMP în fiecare tulpină Finegoldia secvențiată de genom (CAMP1 și CAMP2; figura S4); fiecare omolog adăpostește domeniul familiei factorului CAMP (Pfam07373). Interesant este că un omolog (CAMP2) pe tulpină conține un domeniu suplimentar de tip IG bacterian (Pfam02368). În plus față de CAMP1 și CAMP2, patru ” F. tulpinile „nericia” conțin doi factori CAMP suplimentari care diferă substanțial de ceilalți omologi (CAMP3 și CAMP4; figura S4). Astfel, patru din șapte tulpini „F. nericia” adăpostesc patru gene ale factorului CAMP.
în continuare, am vrut să știm dacă tulpinile Finegoldia produc o reacție CAMP pozitivă. O analiză a plăcii de agar CAMP Arată că unele tulpini sunt puternic pozitive pentru reacția CAMP, în timp ce pentru alte tulpini se poate detecta doar o mică co-hemoliză în condițiile de cultivare aplicate la două și cinci zile de incubație (Fig. 5, Figura S5). Toate cele trei testate F. tulpinile magna au fost puternic pozitive pentru reacția CAMP. Dintre tulpinile „F. nericia” am detectat o oarecare eterogenitate; două tulpini au fost puternic pozitive cu reacție CAMP, patru tulpini au fost doar ușor co-hemolitice, iar o tulpină, CCUG54800, a fost clar negativă cu reacție CAMP. Analiza genomică a constatat că în tulpina CCUG54800 gena care codifică CAMP1 conținea o inserție a unui locus care codifică transpozaza în capătul 5’al genei, care perturbă gena (figura S6).

reacția CAMP a tulpinilor Finegoldia. Au fost utilizate plăci de agar TSA cu 5% sânge de oaie. Staphylococcus aureus este inoculat în banda de mijloc. Un test CAMP pozitiv este indicat prin Liza completă a eritrocitelor la interfața Finegoldia sp. și dungile S. aureus. Toate cele trei tulpini F. magna (în roșu) și două din șapte tulpini „F. nericia” (în albastru) au prezentat o puternică reacție pozitivă CAMP după două zile de incubație anaerobă.
rezultatele noastre indică faptul că factorii CAMP de Finegoldia sp. poate fi funcțional și că există diferențe specifice tulpinii în activitatea factorului CAMP.
două sisteme de teste biochimice nu pot diferenția fără echivoc F. magna de „F. nericia”
reacțiile biochimice ale GPAC inclusiv tulpinile de Finegoldia sp. au fost descrise; doar puține variații biochimice între Finegoldia sp. tulpinile au fost raportate anterior2, 7. Am testat două sisteme de testare biochimice disponibile în comerț care sunt utilizate pentru identificarea anaerobilor. Kitul „API 20A” conține 21 de teste, dintre care 16 teste pentru utilizarea carbohidraților. După cum sa raportat anterior7, niciunul dintre carbohidrați nu a putut fi metabolizat de orice Finegoldia sp. tulpina (datele nu sunt afișate). Celelalte reacții (indol, urează, catalază, hidroliza esculinei și lichefierea gelatinei) nu au evidențiat nici o diferență între tulpinile de F. magna și „F. nericia” (datele nu sunt prezentate). Kitul „RapIDTM ANA II” conține 18 teste pentru activități enzimatice. În timp ce enzimele zaharolitice lipsesc, toate Finegoldia sp. tulpinile produc enzime proteolitice (tabelul S5). Reacția de detectare a activității fosfatazei a fost singura reacție care a diferit între tulpini, toate tulpinile F. magna aparent negative și majoritatea tulpinilor „F. nericia” pozitive. Cu toate acestea, interpretarea acestui rezultat al testului este limitată, deoarece schimbarea culorii a fost la limita semnificației. În concluzie, două sisteme comerciale existente de testare biochimică, API 20 A și RapIDTM ANA II, nu pot diferenția fără echivoc F. magna de”F. nericia”. Proprietățile biochimice ale Finegoldia sp. trebuie să fie explorate în continuare cu teste biochimice suplimentare.
microscopia relevă diferențe de suprafață între F. magna și „F. nericia”
într-o încercare de a vizualiza morfologia și structura suprafeței lui F. magna și „F. nericia ” am folosit microscopie electronică de scanare (SEM) și microscopie de forță atomică (AFM) pe tulpini de F. magna (07T609, 08t492 și 09T408) și „F. nericia” (09t494, 12t272 și 12t306). Sem a detectat diferențe de suprafață: celulele F. magna au fost mai aderente sau agregative, producând aparent o matrice polimerică extracelulară (Fig. 6). Acest lucru confirmă lucrările anterioare care au vizualizat o matrice exopolizaharidică în biofilmele F. magna (tulpina fmbs12) prin microscopia electronică de scanare, precum și microscopia de scanare laser confocală23. În testul ” F. nericia ” tulpini nu am putut identifica o astfel de matrice; celulele par mai puțin agregative.
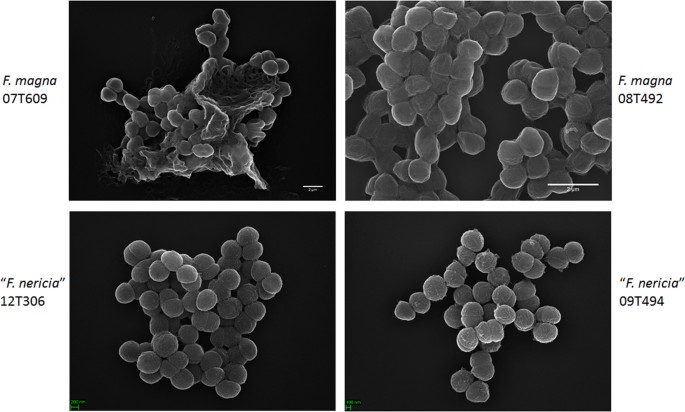
comparația microscopică electronică de scanare a F. magna și”F. nericia”. Panourile superioare și inferioare prezintă imagini reprezentative ale F. magna 07T609 și 08T492 și” F. nericia ” 09T494 și, respectiv, 12T306. Celulele tulpinilor F. magna sunt mai aderente / agregative și par să producă o matrice polimerică, în special tulpina 07T609; în schimb, celulele „F. nericia” sunt mai puțin agregative.
AFM a confirmat aceste observații. Mai mult, am observat diferențe în ceea ce privește apendicele celulare. Celulele F. magna au produs anexe filamentoase mai lungi care ieșeau de pe întreaga suprafață celulară; aceste structuri ar putea reprezenta pili sau fimbrii (Fig. 7). În schimb, celulele tulpinilor „F. nericia” aveau o suprafață mai netedă, fără anexe lungi proeminente. Cu toate acestea, nu se poate exclude faptul că aceste observații AFM depind de condițiile de cultivare bacteriană și de etapele de prelucrare a probelor.

comparație microscopică a Forței Atomice a lui F. magna și”F. nericia”. Panourile din stânga și din dreapta reprezintă imagini ale lui F. magna 07T609 și, respectiv, „F. nericia” 12T306. (A și C) imagini de înaltă rezoluție cu metoda soft-touch avansată asistată, cu bare de scară Z de până la 0,3 mm și bare XY care indică 500 nm. (B și D) suprapunerea imaginilor cu contrast ridicat ale (a și C), combinate cu îndepărtarea polinomială orizontală și verticală independentă a (a și C), rezolvate la gradul 11. Experimentele au fost efectuate pe trei probe din fiecare tulpină.