- Genom av stammar av Finegoldia
- uppdelning av Finegoldia-stammar i två distinkta klader baserat på genomjämförelse
- Stamspecifik heterogenitet av Finegoldia sp
- Värdinteraktionsfaktorer och (förmodade) virulensfaktorer
- Finegoldia genom kodar CAMP-faktorer som är funktionella
- två biokemiska testsystem kan inte entydigt skilja F. magna från ”F. nericia”
- mikroskopi avslöjar ytskillnader mellan F. magna och ”F. nericia”
Genom av stammar av Finegoldia
totalt 17 Genom av Finegoldia sp. analyserades och jämfördes. Tio stammar har nyligen sekvenserats av vår grupp15; de isolerades från patienter med ortopediska gemensamma implantatassocierade infektioner i Sverige. Sju genom sekvenserades tidigare av andra, inklusive det slutna genomet av typstammen ATCC 2932814. Genomfunktioner hos alla hittills sekvenserade stammar av Finegoldia sammanfattas i Tabell 1. Anteckningen med Prokka detekterades mellan 1570 och 1906 kodande sekvenser (CDS) per genom, med i genomsnitt 1760 CDS per stam.
uppdelning av Finegoldia-stammar i två distinkta klader baserat på genomjämförelse
vi märkte tidigare en stor heterogenitet bland Genom av Finegoldia-isolater15. En kärngenominriktning av alla 17 genom gjordes och enstaka nukleotidpolymorfismer (SNP) i kärngenomet kallades. Totalt identifierades 126 647 kärngenom SNP: er och användes för fylogenomisk rekonstruktion, vilket avslöjade att de 17 genomerna kan grupperas i två distinkta klader (Fig. 1). En ytterligare analys som bestämmer den genomsnittliga nukleotididentifieringen (ANI) av kärngenomet visade att de två kladerna uppvisar en ANI på 90,7%, med en stamspecifik ANI-variation från 90,2% till 91,2%. En sådan låg ANI indikerar att de två kladerna representerar enskilda arter16. En clade innehåller mest tidigare sekvenserade och studerade stammar, inklusive ATCC29328 och ALB8; det kan betraktas som den klassiska F. magna clade, med en intraclade ANI på 96,6% (variation 95,1–97,4%). Den andra kladen med en intraclade ANI på 94,8% (93,4–96.6%) innehåller en annan ATCC-stam (ATCC 53516) och huvudsakligen stammar isolerade från patienter vid Universitetssjukhuset i Saudirebro, Sverige15. Hädanefter benämns stammar av denna klad preliminärt ”Finegoldia nericia” (Latin för N Auguirke, den svenska regionen, där de flesta stammar av denna art isolerades).

Fylogenomiskt träd baserat på kärngenom SNP: er av alla 17 sekvenserade genom av Finegoldia-stammar. Programmet Parsnp användes för att anpassa kärngenomet och ringa SNP. Kärngenomet representeras av 43% av referensgenomet (ATCC29328); totalt 126 647 pålitliga kärngenom SNP användes för att rekonstruera en helgenom fylogeni. Genom kan tilldelas två huvudklader; en klad representerar F. magna-stammar och den andra kladen består av stammar av en ny art, här preliminärt benämnd ”F. nericia”.
en jämförelse av 16S rRNA-sekvenser av alla isolat avslöjade en hög grad av total likhet (> 99%), men detekterade en separation av F. magna-isolat från ”F. nericia” – isolat på grund av 5 till 9 ”F. nericia ” – specifika SNP (figur S1). Det avslöjade också en högre diversifiering bland” F. nericia ” – isolat jämfört med F. magna. Därefter analyserade vi ytterligare 16s rRNA-sekvenser som tillhör Finegoldia sp. lagras i GenBank (figur S2). Detta avslöjade att de flesta lagrade sekvenser kluster med” F. nericia”, i synnerhet en grupp av Finegoldia sp. stammar isolerade från gallsten biofilms17. Återigen detekteras en högre diversifiering bland” F. nericia ” – isolat. Därefter analyserade vi humana mikrobiomdata lagrade i IMG-databasen (Integrated Microbial Genomes and microbiome samples) för närvaron av F. magna och ”F. nericia” – sekvenser. 3287 och 7068 kodande sekvenser av F. magna ATCC29328 respektive” F. nericia ” ATCC 53516 kunde identifieras i de 875 sammansatta humana mikrobiomdatauppsättningarna som finns i IMG (data visas inte); högsta förekomster av Finegoldia detekterades i retroaurikulära veckprover. Sammantaget tyder dessa analyser på att bland humanassocierade Finegoldiaisolat ”F. nericia” verkar vara vanligare än F. magna.
Stamspecifik heterogenitet av Finegoldia sp
för att ytterligare belysa befolkningsstrukturen i finegoldia-släktet utfördes jämförande analyser av 12 genom (alla tio nyligen sekvenserade stammar och de två ATCC-stammarna), inklusive fyra stammar av F. magna-kladen och åtta stammar av ”F. nericia” – kladen. En visuell jämförelse av de 12 genomerna markerade nukleotididentitetsskillnaden mellan de två kladerna (Fig. 2). Intressant nog verkar plasmid som finns i ATCC 29328 vara specifik för denna stam eftersom ingen annan sekvenserad stam innehåller en liknande sekvens (Fig. 2A).

genom jämförelse av 12 stammar som tillhör släktet Finegoldia. De två innersta ringarna representerar G + C-innehållet (svart) och GC-skev (violett/grön). (A) referensstammen är ATCC29328 (F. magna); notera den höga nukleotididentiteten (visualiserad av den mörkröda färgen) av de tre innersta genomerna (som tillhör alla F. magna), och den nedre identifiera åtta stammar som tillhör ”F. nericia” (blek röd färg på de åtta yttre ringarna); från insidan till utsidan: stammar 07T609, 08T492, 09t408, 09t494, 12t272, 12t273, 12t306, CCUG 54800, T151023, T160124 och ATCC 53516. Den stora ATCC29328-specifika regionen i den övre vänstra regionen representerar plasmid pFMC. (B) referensstammen är 09T494 (”F. nericia”). De sju” F. nericia ” – genomerna (inre ringar) har en högre nukleotid identifiera (representerad av den mörkare röda färgen), jämfört med de fyra F. magna-genomerna( yttre ringar); från insidan till utsidan: stammar 12T272, ATCC 53516, 12t306, 12t273, CCUG 54800, T151023, T160124, ATCC29328, 07t609, 08T492 och 09T408. Det finns inga tecken på”F. nericia” -specifika genomiska regioner, men notera de fem stora stamspecifika klusterna.
inga signifikanta skillnader i genomstorlek och antal CD-skivor detekterades mellan de två kladerna. Dessutom detekterades inga clade-specifika (dvs F. magna eller ”F. nericia”-specifika) genomiska regioner, som bedömdes från BRIG-analysen (Fig. 2) och bekräftas också av en dubbelriktad sprängmetod för alla CD-skivor med ProteinOrtho (tabell S1). Detta tillvägagångssätt upptäckte 1202 orthologs delade av alla 12 genom, dvs i genomsnitt 68% av CD-skivorna för varje stam är en del av kärnproteomen. Endast mycket få kladspecifika gener finns. Istället kan många regioner som delas av endast ett fåtal stammar hittas, t.ex. för stam 09T494 (se även Fig. 2B). Dessutom finns stamspecifika regioner som kodar för totalt 1016 stamspecifika CD-skivor, med ett stort intervall mellan stammarna, t.ex. 187 och 22 stamspecifika CD-skivor detekteras i stam ATCC29328 respektive T151023 (tabell S1). Bland de stamspecifika funktionerna finns olika transportfunktioner (järn, effluxpumpar, oligopeptid ABC-transportörer), ytproteiner, restriktionsmodifieringssystem, konjugativa överföringsfunktioner, polyketidsyntes, bakteriociner, fagrelaterade funktioner, antibiotikaresistensbestämningsfaktorer och CRISPR/cas-system.
intressant nog upptäcktes skillnader i CRISPR/cas-systemen mellan stammar av F. magna och ”F. nericia” (tabell S2). Sju av nio F. magna-stammar innehöll en till fyra CRISPR-arrayer per stam, innehållande i genomsnitt 12 distanser (från 3 till 38 distanser per stam). Upprepningen” GTTTGAGAATGATGTAATTCATATAGGTATTAAAC ” var specifik för stammar av F. magna. Däremot Bar” F. nericia ” – stammar endast en CRISPR-array per stam som innehöll i genomsnitt 53 distanser (intervall 14 till 87 per stam). Varje stam innehöll en individuell uppsättning distanser i deras CRISPR-array( er), underliggande stamspecifik heterogenitet, och indikerar att varje stam har en unik evolutionär historia.
för att utvärdera om sådana stamspecifika funktioner förvärvades horisontellt tittade vi på tecken på deras rörlighet/förvärv med hjälp av IslandViewer-programmet (figur S3). Detta program förutspådde 141 (stam 09T408) till 408 (stam 12t273) gener belägna i genomiska öar som sannolikt kommer att förvärvas horisontellt, inklusive många av de ovan nämnda stamspecifika generna som är involverade i specialisering, fitness, överlevnad och interspecies konkurrens (tabell S3).
Värdinteraktionsfaktorer och (förmodade) virulensfaktorer
flera värdinteraktionsfaktorer har beskrivits för F. magna 4,5. Sortasberoende pili har identifierats som är viktiga för vidhäftning och kolonisering13. Ett genomiskt locus som kodar för generna för pilus-subenheten Fmp1, den förmodade subenheten Fmp2 och tre sortaser bevaras bland alla sekvenserade genom (Fig. 3A). Det finns stamspecifika variationer upp-och nedströms denna plats, dvs. vissa stammar har ytterligare gener uppströms fmp1 och nedströms om sortas locus, inklusive en gen som kodar för ett hemolysin III-familjeprotein. Intressant nog finns det stamspecifika skillnader angående fmp1. Nästan varje stam bär en individuell variant och en fylogenetisk analys av Fmp1 på proteinnivå visade ingen korrelation med fylogenomisk analys av kärngenomet (Fig. 3B). Däremot är Fmp2 mycket bevarad bland alla stammar. Ett annat protein, kodat uppströms fmp1, har kollagen – och fibrinogenbindande domäner; den kollagenbindande domänen (COG4932) finns också i Fmp2. Vi föreslår att detta protein också tillhör pilus locus och namnger motsvarande gen fmp3.

Sortasberoende pili-kodande genomiska loci i genomerna av Finegoldia-stammar. (A) genomisk organisation av loci i F. magna (röd) och ”F. nericia” (ljusblå) stammar. De tre sortaser generna avbildas i ljusgrön (märkt ”2”); genen för den stora pilus-underenheten Fmp1 visas i rött (märkt ”1”) och fmp2 avbildas i orange (märkt ”3”). Uppströms fmp1 och nedströms sortasgenerna i vissa stammar finns ytterligare gener (märkta ”8”, hemolysin III-familjeprotein och ”10”, hypotetiskt protein) närvarande. Ett förmodat kollagenbindande protein, Fmp3 (märkt ”4”), kodas uppströms fmp1. (B) fylogenetiskt träd av Fmp1 av Finegoldia-stammar. De flesta stammar har en individuell fmp1-variant. Den evolutionära historien härleddes genom att använda maximal Sannolikhetsmetod, gjord i MEGA7.
Protein L är förmodligen den bäst studerade faktorn för F. magna 8. Det är en superantigen som riktar sig mot B-celler och immunoglobuliner. Det har detekterats i cirka 10% av de hittills testade Finegoldia-isolaten, men varierar i storlek och antal antigenbindande fragment (Fab)-bindande domäner (B1-domän, pfam02246)4,8,18. En sprängsökning med det studerade proteinet L från stam 312 (locus ID: M86697, 719 aa) avslöjar några mycket liknande (proteinidentitet över 50%) proteiner med varierande storlekar (603 till 1417 aa) i några stammar, dvs. stammar CCUG54800, T161024, ATCC53516, 12t273 och 12t306 (Fig. 4A). Alla dessa är” F. nericia ” -stammar, vilket indikerar att B-cellens superantigen är vanligare hos denna art. Den exceptionellt stora (1417 aa) protein L-liknande faktorn i stam 12t306 har förutom två B1-domäner också två GA-moduler som är kända från peptostreptococcal albuminbindande proteiner (se nedan). Många ytterligare proteiner med låg likhet med protein L, men utan att innehålla några uppenbara B1-domäner kodas i de genomsekvenserade Finegoldia-stammarna (tabell S4A); återigen finns stamspecifika variationer, och några stammar, dvs 09T408, 09t494 och T151023 har inga sådana homologer.

genomisk loci kodar protein L och FAF homologer i Finegoldia stammar. (A) Sprängsökningar med det karakteriserade proteinet L från stam 312 (locus ID: M86697) utfördes i genomsekvenserade Finegoldiastammar, och homologer (i rött, märkt ”1”) med hög likhet (>50% proteinidentitet) visas (Se även tabell S4A). Endast” F. nericia ” stammar men inte F. magna-stammar innehåller sådana homologer med varierande längder. (B) Blast-sökningar med det studerade FAF-proteinet av stam ALB8 hittade homologer i full längd (i rött, märkt ”1”) i tre F. magna (röd) och fyra ”F. nericia” (ljusblå) stammar. FAF-genen är frameshifted i de två ”F. nericia” – stammarna T151023 och 12t272.
Finegoldia magna befanns binda till humant albumin9. Ett peptostreptokockalbuminbindande protein (PAB) har studerats i stammen ALB8; dess aktivitet är kopplad till en accelererad tillväxthastighet9. PAB-proteinet av stam ALB8 (locus ID: X77864) innehåller GA-moduler, den albuminbindande domänen; strukturen för GA-modulen har lösts upp19. En Sprängsökning avslöjade att en nära PAB-homolog inte kan hittas i andra sekvenserade Finegoldia-genom (tabell S4B). Istället finns det flera homologer som delar upp till 50% proteinidentitet; likheterna mellan homologer i F. magna-stammar är i genomsnitt högre jämfört med homologer i ”F. nericia” – stammar. När det gäller ga-modulinnehållande proteiner kodas totalt 16 proteiner i de 12 Finegoldia-genomerna, vilket indikerar viss redundans av albuminbindande proteiner (tabell S4C). De flesta stammar kodar två proteiner med GA-moduler, förutom från stam 09T408 (F. magna) och T151023 (”F. nericia”).
det cellväggsbundna subtilisinliknande proteaset SufA har visat sig försämra flera humana proteiner, såsom kollagen IV,fibrinogen och antimikrobiella peptider såsom LL-37 och mig/CXCL910,20, 21. Blast-sökningar bestämde att de flesta Finegoldia-stammar bär en SufA-homolog som är mycket lik (proteinidentitet > 75%) till den studerade SufA från stam ALB8 (locus ID: DQ679960). Undantag är stammarna 07T609, 09t408 och CCUG54800. En andra SufA-homolog finns i de flesta stammar; den har en genomsnittlig proteinidentitet på 26% (tabell S4D).
en annan viktig värdinteraktiv faktor är FAF (F. magna adhesion factor). Cirka 90% av F. magna-stammarna producerar denna faktor, som är ansvarig för klumpning av bakterier och förmedlar bindning till källarmembranet genom bindning till BM-4011. Vi fann att sju (tre F. magna och fyra ”F. nericia” – stammar) av de 12 stammarna kodar för en FAF-homolog som liknar den som studerades i ALB8-stammen (Fig. 4B). Strain-specifika skillnader finns och frameshift mutationer ses i” F. nericia ” stammar T151023 och 12t272. Ingen homolog finns i stammarna 09T408, 09t494 och 12t306 (tabell S4E).
Finegoldia genom kodar CAMP-faktorer som är funktionella
vi sökte genomet efter andra förmodade värdinteraktiva faktorer och hittade gener som kodar för Christie-Atkins-Munch-Petersen (CAMP) – faktorer. CAMP-faktorer kan fungera som co-hemolysiner och redogöra för den så kallade CAMP-reaktionen, den synergistiska lysen av får-erytrocyter av Staphylococcus aureus-sfingomyelinas C (beta-toxin) och en CAMP-faktor22.
genombrytning identifierade två gener som kodar för CAMP-faktorer i varje genomsekvenserad Finegoldia-stam (CAMP1 och CAMP2; figur S4); varje homolog har CAMP factor family-domänen (Pfam07373). Intressant nog har en homolog (CAMP2) per stam en ytterligare bakteriell ig-liknande domän (Pfam02368). Förutom CAMP1 och CAMP2, fyra ” F. nericia ” – stammar innehåller ytterligare två CAMP-faktorer som väsentligt skiljer sig från de andra homologerna (CAMP3 och CAMP4; figur S4). Således har fyra av sju ”F. nericia” – stammar fyra CAMPFAKTORGENER.
därefter ville vi veta om Finegoldia-stammar ger en positiv CAMP-reaktion. En CAMP-agarplattanalys visar att vissa stammar är starkt CAMP-reaktionspositiva medan för andra stammar endast liten co-hemolys kan detekteras under de applicerade odlingsförhållandena vid två och fem dagars inkubation (Fig. 5, Figur S5). Alla tre testade F. magna-stammar var starkt LÄGERREAKTIONSPOSITIVA. Bland” F. nericia ” – stammar upptäckte vi viss heterogenitet; två stammar var starkt CAMP-reaktionspositiva, fyra stammar var bara lite co-hemolytiska och en stam, CCUG54800, var tydligt CAMP-reaktion-negativ. Genomisk analys fann att i stam CCUG54800 innehöll CAMP1-kodande genen en insättning av ett transposas-kodande locus i 5′-änden av genen, vilket stör genen (figur S6).

CAMP reaktion av Finegoldia stammar. Tsa-agarplattor med 5% fårblod användes. Staphylococcus aureus ympas i mitten. Ett positivt CAMP-test indikeras av fullständig erytrocytlys vid gränssnittet för Finegoldia sp. och S. aureus strimmor. Alla tre F. magna (i rött) stammar och två av sju ”F. nericia” (i blått) stammar visade en stark positiv CAMP-reaktion efter två dagars anaerob inkubation.
våra resultat tyder på att CAMP faktorer av Finegoldia sp. kan vara funktionell och att det finns töjningsspecifika skillnader i CAMP-faktoraktivitet.
två biokemiska testsystem kan inte entydigt skilja F. magna från ”F. nericia”
biokemiska reaktioner av GPAC inklusive stammar av Finegoldia sp. har beskrivits; endast liten biokemisk variation bland Finegoldia sp. stammar var tidigare rapporterade2, 7. Vi testade två kommersiellt tillgängliga biokemiska testsystem som används för identifiering av anaerober. Satsen” API 20A ” innehåller 21 tester, bland annat 16 tester för kolhydratutnyttjande. Som rapporterats tidigare7 kunde ingen av kolhydraterna metaboliseras av någon Finegoldia sp. stam (data visas inte). De andra reaktionerna (indol, ureas, katalas, esculinhydrolys och gelatinvätskning) avslöjade inte heller någon skillnad mellan stammar av F. magna och ”F. nericia” (data visas inte). Satsen” RapIDTM ANA II ” innehåller 18 test för enzymatiska aktiviteter. Medan sackarolytiska enzymer saknas, alla Finegoldia sp. stammar producerar proteolytiska enzymer (tabell S5). Reaktionen som detekterade fosfatasaktivitet var den enda reaktionen som skilde sig mellan stammarna, med alla F. magna-stammar tydligen negativa och de flesta ”F. nericia” – stammar positiva. Tolkningen av detta testresultat är dock begränsad eftersom färgförändringen var på gränsen till Betydelse. Sammanfattningsvis kan två befintliga kommersiella biokemiska testsystem, API 20 A och RapIDTM ANA II, inte entydigt skilja F. magna från ”F. nericia”. De biokemiska egenskaperna hos Finegoldia sp. behöver undersökas ytterligare med ytterligare biokemiska tester.
mikroskopi avslöjar ytskillnader mellan F. magna och ”F. nericia”
i ett försök att visualisera morfologin och ytstrukturen hos F. magna och ”F. nericia ” vi använde scanningelektronmikroskopi (SEM) och atomkraftmikroskopi (AFM) på stammar av F. magna (07T609, 08t492 och 09T408) och ”F. nericia” (09T494, 12t272 och 12t306). SEM detekterade ytskillnader: celler av F. magna var mer vidhäftande eller aggregerande, till synes producerar en extracellulär polymermatris (Fig. 6). Detta bekräftar tidigare arbete som har visualiserat en exopolysackaridmatris i biofilmer av F. magna (stam FmBs12) genom scanningelektronmikroskopi såväl som konfokal laserskanningsmikroskopi23. I den testade ” F. nericia ” stammar vi kunde inte identifiera en sådan matris; celler verkar mindre aggregerande.
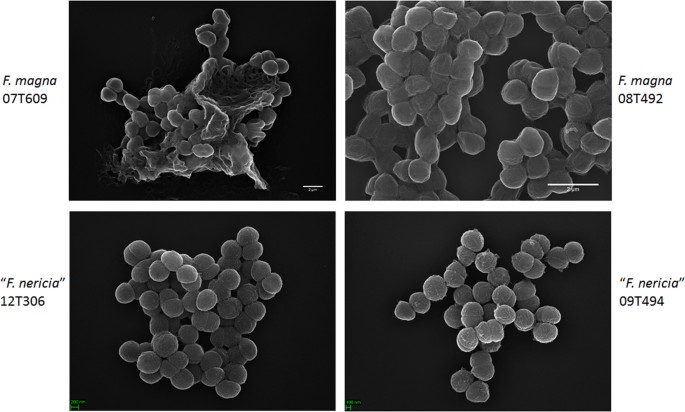
Scanningelektronmikroskopisk jämförelse av F. magna och”F. nericia”. De övre och nedre panelerna visar representativa bilder av F. magna 07t609 respektive 08T492 respektive ”F. nericia” 09T494 respektive 12t306. Celler av F. magna-stammar är mer vidhäftande / aggregerande och verkar producera en polymermatris, i synnerhet stam 07T609; däremot är celler av ”F. nericia” mindre aggregerande.
AFM bekräftade dessa observationer. Dessutom observerade vi skillnader när det gäller cellbilagor. Celler av F. magna producerade längre filamentösa bilagor som sticker ut från hela cellytan; dessa strukturer kan representera pili eller fimbriae (Fig. 7). Däremot hade celler av” F. nericia ” – stammar en jämnare yta utan långa utskjutande bilagor. Det kan emellertid inte uteslutas att dessa AFM-observationer beror på bakterieodlingsförhållandena och på provbehandlingsstegen.

Atomkraft mikroskopisk jämförelse av F. magna och”F. nericia”. Vänster och höger paneler representerar bilder av F. magna 07t609 respektive” F. nericia ” 12t306. (A och C) högupplösta bilder med assisterad avancerad soft-touch-metod med Z-skalstänger upp till 0,3 oz.m och XY-staplar som indikerar 500 nm. (B och D) Overlay bilder av hög kontrast bilder av (A och C), i kombination med oberoende horisontella och vertikala polynom avlägsnande av (A och C), löst till den 11: e graden. Experiment utfördes på tre prover från varje stam.