- Génomes de souches de Finegoldia
- Division des souches de Finegoldia en deux clades distincts basés sur la comparaison du génome
- Hétérogénéité spécifique à la souche de Finegoldia sp
- Facteurs interagissant avec l’hôte et facteurs de virulence (putatifs)
- Les génomes de Finegoldia codent des facteurs de l’AMPC fonctionnels
- Deux systèmes de tests biochimiques ne peuvent pas différencier sans ambiguïté F. magna de « F. nericia »
- La microscopie révèle des différences de surface entre F. magna et « F. nericia »
Génomes de souches de Finegoldia
Au total 17 génomes de Finegoldia sp. ont été analysés et comparés. Dix souches ont récemment été séquencées par notre groupe15; elles ont été isolées chez des patients atteints d’infections articulaires orthopédiques associées à des implants en Suède. Sept génomes ont été précédemment séquencés par d’autres, y compris le génome fermé de la souche type ATCC 2932814. Les caractéristiques du génome de toutes les souches séquencées de Finegoldia jusqu’à présent sont résumées dans le tableau 1. L’annotation utilisant Prokka a détecté entre 1570 et 1906 séquences codantes (CD) par génome, avec une moyenne de 1760 CD par souche.
Division des souches de Finegoldia en deux clades distincts basés sur la comparaison du génome
Nous avons précédemment remarqué une grande hétérogénéité entre les génomes des isolats de Finegoldia 15. Un alignement noyau-génome des 17 génomes a été effectué et des polymorphismes nucléotidiques simples (SNP) dans le génome noyau ont été appelés. Au total, 126 647 SNP du génome central ont été identifiés et utilisés pour la reconstruction phylogénomique, révélant que les 17 génomes peuvent être regroupés en deux clades distincts (Fig. 1). Une analyse supplémentaire qui détermine l’identification nucléotidique moyenne (ANI) du génome du noyau a montré que les deux clades présentent un ANI de 90,7%, avec une variation d’ANI spécifique à la souche de 90,2% à 91,2%. Un ANI aussi bas indique que les deux clades représentent des espèces individuelles16. Un clade contient la plupart des souches précédemment séquencées et étudiées, y compris ATCC29328 et ALB8; il peut être considéré comme le clade classique de F. magna, avec un ANI intraclade de 96,6% (variation de 95,1 à 97,4%). L’autre clade avec un ANI intraclade de 94,8% (93,4-96.6%) contient une autre souche ATCC (ATCC 53516) et principalement des souches isolées de patients de l’Hôpital universitaire d’Örebro, en Suède15. Ci-après, les souches de ce clade sont provisoirement nommées « Finegoldia nericia » (en latin pour Närke, la région suédoise, où la plupart des souches de cette espèce ont été isolées).

Arbre phylogénomique basé sur les SNP du génome central des 17 génomes séquencés des souches de Finegoldia. Le programme Panais a été utilisé pour aligner le génome du noyau et appeler des SNP. Le génome central est représenté par 43% du génome de référence (ATCC29328); un total de 126 647 SNP fiables du génome central ont été utilisés pour reconstruire une phylogénie du génome entier. Les génomes peuvent être affectés à deux clades principaux ; un clade représente des souches de F. magna et l’autre clade est constitué de souches d’une nouvelle espèce, ici provisoirement nommée « F. nericia ».
Une comparaison des séquences d’ARNr 16S de tous les isolats a révélé un degré élevé de similitude globale (> 99%), mais a détecté une séparation des isolats de F. magna des isolats de « F. nericia », en raison de 5 à 9″F. nericia » – SNP spécifiques (Figure S1). Il a également révélé une plus grande diversification parmi les isolats de « F. nericia » par rapport à F. magna. Ensuite, nous avons analysé d’autres séquences d’ARNr 16S appartenant à Finegoldia sp. stocké dans GenBank (figure S2). Ceci a révélé que la plupart des séquences stockées se regroupent avec « F. nericia », en particulier un groupe de Finegoldia sp. souches isolées des biofilms du stent biliaire17. Encore une fois, une plus grande diversification parmi les isolats de « F. nericia » est détectée. Ensuite, nous avons analysé les données du microbiome humain stockées dans la base de données IMG (Integrated Microbian Genomes and microbiome samples) pour détecter la présence de séquences de F. magna et de « F. nericia ». 3287 et 7068 séquences codantes de F. magna ATCC29328 et de « F. nericia » ATCC 53516, respectivement, ont pu être identifiées dans les 875 ensembles de données de microbiome humain assemblés présents dans l’IMG (données non illustrées); les occurrences les plus élevées de Finegoldia ont été détectées dans des échantillons de plis rétroauriculaires. Prises ensemble, ces analyses suggèrent que parmi les isolats de Finegoldia associés à l’homme, « F. nericia » semble être plus répandu que F. magna.
Hétérogénéité spécifique à la souche de Finegoldia sp
Pour mieux éclairer la structure des populations du genre Finegoldia, des analyses comparatives de 12 génomes (les dix souches nouvellement séquencées et les deux souches ATCC) ont été effectuées, dont quatre souches du clade F. magna et huit souches du clade « F. nericia ». Une comparaison visuelle des 12 génomes a mis en évidence la différence d’identité nucléotidique entre les deux clades (Fig. 2). Fait intéressant, le plasmide trouvé dans ATCC 29328 semble être spécifique de cette souche car aucune autre souche séquencée ne contient une séquence similaire (Fig. 2 BIS).

Comparaison du génome de 12 souches appartenant au genre Finegoldia. Les deux anneaux les plus à l’intérieur représentent la teneur en G+ C (noir) et l’inclinaison GC (violet/vert). (A) La souche de référence est ATCC29328 (F. magna) ; notez l’identité nucléotidique élevée (visualisée par la couleur rouge foncé) des trois génomes les plus internes (appartenant tous à F. magna), et la partie inférieure identifient à huit souches appartenant à « F. nericia » (couleur rouge pâle des huit anneaux externes); de l’intérieur vers l’extérieur : souches 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 et ATCC 53516. La grande région spécifique à ATCC29328 dans la région supérieure gauche représente le plasmide pFMC. B) La souche de référence est 09T494 (« F. nericia »). Les sept génomes de « F. nericia » (anneaux internes) ont une identification nucléotidique plus élevée (représentée par la couleur rouge plus foncée), par rapport aux quatre génomes de F. magna (anneaux externes); de l’intérieur vers l’extérieur: souches 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 et 09T408. Il n’y a aucun signe de régions génomiques spécifiques à « F. nericia », mais notez les cinq grands groupes spécifiques à la souche.
Aucune différence significative dans la taille du génome et le nombre de CD n’a été détectée entre les deux clades. De plus, aucune région génomique spécifique à un clade (c’est-à-dire spécifique à F. magna ou à » F. nericia « ) n’a été détectée, comme en témoigne l’analyse de BRIG (Fig. 2) et également confirmé par une approche de blast bidirectionnelle de tous les CD utilisant ProteinOrtho (tableau S1). Cette approche a permis de détecter 1202 orthologues partagés par les 12 génomes, c’est-à-dire qu’en moyenne 68% des CD de chaque souche font partie du protéome central. On ne trouve que très peu de gènes spécifiques à un clade. Au lieu de cela, de nombreuses régions partagées par seulement quelques souches peuvent être trouvées, par exemple pour la souche 09T494 (voir également Fig. 2B). En outre, des régions spécifiques à la souche sont présentes qui codent au total 1016 CD spécifiques à la souche, avec une large gamme entre les souches, par exemple 187 et 22 CD spécifiques à la souche sont détectés dans les souches ATCC29328 et T151023, respectivement (tableau S1). Parmi les fonctions spécifiques à la souche figurent diverses fonctions de transport (fer, pompes d’efflux, transporteurs d’oligopeptides ABC), protéines de surface, systèmes de modification de restriction, fonctions de transfert conjugatif, synthèse de polykétides, bactériocines, fonctions liées aux phages, déterminants de la résistance aux antibiotiques et systèmes CRISPR / cas.
Il est intéressant de noter que des différences dans les systèmes CRISPR/cas ont été détectées entre les souches de F. magna et de » F. nericia » (tableau S2). Sept F sur neuf. les souches magna contenaient un à quatre réseaux CRISPR par souche, contenant en moyenne 12 espaceurs (allant de 3 à 38 espaceurs par souche). La répétition » GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC » était spécifique aux souches de F. magna. En revanche, les souches de « F. nericia » ne portaient qu’un réseau CRISPR par souche qui contenait en moyenne 53 espaceurs (plage de 14 à 87 par souche). Chaque souche contenait un ensemble individuel d’espaceurs dans leur(s) réseau(s) CRISPR, ce qui sous-tend l’hétérogénéité spécifique à la souche et indique que chaque souche a une histoire évolutive unique.
Afin d’évaluer si de telles fonctions spécifiques à la souche ont été acquises horizontalement, nous avons examiné les signes de leur mobilité / acquisition à l’aide du programme IslandViewer (Figure S3). Ce programme a prédit 141 gènes (souche 09T408) à 408 gènes (souche 12T273) situés dans des îles génomiques susceptibles d’être acquis horizontalement, y compris un grand nombre des gènes spécifiques à la souche susmentionnés impliqués dans la spécialisation, la condition physique, la survie et la compétition interspécifique (tableau S3).
Facteurs interagissant avec l’hôte et facteurs de virulence (putatifs)
Plusieurs facteurs interagissant avec l’hôte ont été décrits pour F. magna 4,5. On a identifié des pili dépendants de la sortase qui sont importants pour l’adhésion et la colonisation13. Un locus génomique qui code les gènes de la sous-unité du pilus Fmp1, de la sous-unité putative Fmp2 et de trois sortases est conservé parmi tous les génomes séquencés (Fig. 3 BIS). Il existe des variations spécifiques à la déformation en amont et en aval de ce locus, c’est-à-dire certaines souches possèdent des gènes supplémentaires en amont de fmp1 et en aval du locus de la sortase, y compris un gène codant pour une protéine de la famille de l’hémolysine III. Fait intéressant, il existe des différences spécifiques à la souche en ce qui concerne le fmp1. Presque chaque souche porte un variant individuel et une analyse phylogénétique de Fmp1 au niveau de la protéine n’a montré aucune corrélation avec l’analyse phylogénétique du génome du noyau (Fig. 3B). En revanche, le Fmp2 est très conservé parmi toutes les souches. Une autre protéine, codée en amont de fmp1, abrite des domaines de liaison au collagène et au fibrinogène; le domaine de liaison au collagène (COG4932) est également présent dans Fmp2. Nous proposons que cette protéine appartienne également au locus pilus et nommons le gène correspondant fmp3.

Loci génomiques codant pour le pili dépendant de la sortase dans les génomes des souches de Finegoldia. (A) Organisation génomique des loci chez les souches de F. magna (rouge) et de « F. nericia » (bleu clair). Les trois gènes sortases sont représentés en vert clair (étiquetés « 2 »); le gène de la sous-unité principale du pilus Fmp1 est représenté en rouge (marqué « 1 ») et fmp2 est représenté en orange (marqué « 3 »). En amont de la fmp1 et en aval des gènes de la sortase dans certaines souches, des gènes supplémentaires (marqués « 8 », protéine de la famille de l’hémolysine III et « 10 », protéine hypothétique) sont présents. Une protéine supposée se liant au collagène, Fmp3 (marquée « 4 »), est codée en amont de fmp1. (B) Arbre phylogénétique de Fmp1 des souches de Finegoldia. La plupart des souches possèdent une variante Fmp1 individuelle. L’histoire évolutive a été déduite en utilisant la méthode du maximum de vraisemblance, effectuée dans MEGA7.
La protéine L est probablement le facteur le mieux étudié de F. magna 8. C’est un superantigène qui cible les cellules B et les immunoglobulines. Il a été détecté dans environ 10% des isolats de Finegoldia testés jusqu’à présent, mais varie en taille et en nombre de domaines de liaison aux fragments de liaison à l’antigène (Fab) (domaine B1, pfam02246) 4,8,18. Une recherche blast avec la protéine L étudiée de la souche 312 (locus ID: M86697, 719 aa) révèle quelques protéines très similaires (identité protéique supérieure à 50%) de tailles variables (603 à 1417 aa) dans quelques souches, i.e. souches CCUG54800, T161024, ATCC53516, 12T273 et 12T306 (Fig. 4 BIS). Toutes ces souches sont des souches de « F. nericia », ce qui indique que le superantigène à cellules B est plus répandu chez cette espèce. Le facteur de type L de la protéine (1417 aa) exceptionnellement grand de la souche 12T306 possède en plus de deux domaines B1 également deux modules GA connus à partir de protéines de liaison à l’albumine peptostreptococcique (voir ci-dessous). De nombreuses protéines supplémentaires ayant une faible similitude avec la protéine L, mais ne contenant aucun domaine B1 apparent, sont codées dans les souches de Finegoldia séquencées par génome (tableau S4A); là encore, des variations spécifiques à la souche existent, et quelques souches, c’est-à-dire 09T408, 09T494 et T151023 ne possèdent pas de tels homologues.

Loci génomiques codant pour les homologues de la protéine L et de la FAF chez les souches de Finegoldia. (A) Des recherches par Blast avec la protéine L caractérisée de la souche 312 (locus ID: M86697) ont été effectuées dans des souches de Finegoldia séquencées dans le génome, et des homologues (en rouge, marqués « 1 ») présentant une forte similitude (> 50% d’identité protéique) sont présentés (voir également le tableau S4A). Seulement les souches « F. nericia » mais pas F. les souches magna contiennent de tels homologues de longueurs variables. (B) Des recherches par Blast avec la protéine FAF étudiée de la souche ALB8 ont trouvé des homologues de pleine longueur (en rouge, étiquetés « 1 ») dans trois souches de F. magna (rouge) et quatre souches de « F. nericia » (bleu clair). Le gène FAF est modifié dans les deux souches de « F. nericia » T151023 et 12T272.
On a constaté que Finegoldia magna se liait à l’albumine9 humaine. Une protéine de liaison à l’albumine peptostreptococcique (PAB) a été étudiée chez la souche ALB8; son activité est liée à un taux de croissance accéléré9. La protéine PAB de la souche ALB8 (locus ID: X77864) contient des modules GA, le domaine de liaison à l’albumine ; la structure du module GA a été résolue19. Une recherche par Blast a révélé qu’un homologue PAB proche ne peut pas être trouvé dans d’autres génomes de Finegoldia séquencés (tableau S4B). Au lieu de cela, il existe plusieurs homologues qui partagent jusqu’à 50% d’identité protéique; les similitudes entre les homologues des souches de F. magna sont en moyenne plus élevées que celles des homologues des souches de « F. nericia ». En ce qui concerne les protéines contenant le module GA, au total 16 protéines sont codées dans les 12 génomes de Finegoldia, ce qui indique une certaine redondance des protéines se liant à l’albumine (tableau S4C). La plupart des souches codent deux protéines avec des modules GA, à l’exception des souches 09T408 (F. magna) et T151023 (« F. nericia »).
La protéase de type subtilisine fixée à la paroi cellulaire SufA dégrade plusieurs protéines humaines, telles que le collagène IV, le fibrinogène et des peptides antimicrobiens tels que LL-37 et MIG/ CXCL910, 20, 21. Les recherches effectuées par Blast ont permis de déterminer que la plupart des souches de Finegoldia portent un homologue de l’ASE très similaire (identité protéique > 75 %) à l’ASE étudiée de la souche ALB8 (locus ID: DQ679960). Les exceptions sont les souches 07T609, 09T408 et CCUG54800. Un deuxième homologue SufA est présent dans la plupart des souches; il a une identité protéique moyenne de 26% (tableau S4D).
Un autre facteur important d’interaction avec l’hôte est le FAF (facteur d’adhérence de F. magna). Environ 90% des souches de F. magna produisent ce facteur, qui est responsable de l’agglutination des bactéries et médie la liaison à la membrane basale en se liant à BM-4011. Nous avons constaté que sept (trois souches de F. magna et quatre souches de « F. nericia ») sur les 12 souches codent un homologue FAF similaire à celui étudié dans la souche ALB8 (Fig. 4B). Des différences spécifiques à la souche existent et des mutations à décalage de cadre sont observées chez les souches T151023 et 12T272 de « F. nericia « . Aucun homologue ne peut être trouvé dans les souches 09T408, 09T494 et 12T306 (tableau S4E).
Les génomes de Finegoldia codent des facteurs de l’AMPC fonctionnels
Nous avons cherché dans le génome d’autres facteurs susceptibles d’interagir avec l’hôte et avons trouvé des gènes codant pour les facteurs Christie-Atkins-Munch-Petersen (AMPC). Les facteurs d’AMPC peuvent agir comme des co-hémolysines et expliquer la réaction dite d’AMPC, la lyse synergique des érythrocytes de mouton par la sphingomyélinase C de Staphylococcus aureus (bêta-toxine) et un facteur d’AMPC22.
L’exploration génomique a permis d’identifier deux gènes codant pour les facteurs AMPC dans chaque souche de Finegoldia séquencée par génome (CAMP1 et CAMP2; Figure S4) ; chaque homologue abrite le domaine familial des facteurs AMPC (Pfam07373). Fait intéressant, un homologue (CAMP2) par souche abrite un domaine bactérien supplémentaire de type Ig (Pfam02368). En plus du CAMP1 et du CAMP2, quatre « F. les souches » nericia » contiennent deux facteurs AMPC supplémentaires qui diffèrent sensiblement des autres homologues (CAMP3 et CAMP4; Figure S4). Ainsi, quatre souches de » F. nericia » sur sept possèdent quatre gènes de facteur AMPC.
Ensuite, nous voulions savoir si les souches de Finegoldia produisaient une réaction d’AMPC positive. Un test sur plaque de gélose de l’AMPC montre que certaines souches sont fortement positives à la réaction de l’AMPC alors que pour d’autres souches, seule une faible co-hémolyse peut être détectée dans les conditions de culture appliquées à deux et cinq jours d’incubation (Fig. 5, Figure S5). Tous les trois testés F. les souches de magna étaient fortement positives à la réaction de l’AMPC. Parmi les souches de « F. nericia », nous avons détecté une certaine hétérogénéité; deux souches étaient fortement positives à la réaction de l’AMPC, quatre souches n’étaient que légèrement co-hémolytiques et une souche, CCUG54800, était clairement négative à la réaction de l’AMPC. L’analyse génomique a révélé que dans la souche CCUG54800, le gène codant pour la CAMP1 contenait une insertion d’un locus codant pour la transposase à l’extrémité 5′ du gène, ce qui perturbe le gène (Figure S6).

Réaction de CAMP des souches de Finegoldia. Des plaques de gélose TSA contenant 5% de sang de mouton ont été utilisées. Staphylococcus aureus est inoculé dans la strie médiane. Un test d’AMPC positif est indiqué par une lyse complète des érythrocytes à l’interface du Finegoldia sp. et les stries de S. aureus. Les trois souches de F. magna (en rouge) et deux des sept souches de « F. nericia » (en bleu) ont montré une forte réaction d’AMPC positive après deux jours d’incubation anaérobie.
Nos résultats indiquent que les facteurs d’AMPC de Finegoldia sp. peut être fonctionnel et qu’il existe des différences spécifiques à la souche dans l’activité du facteur AMPC.
Deux systèmes de tests biochimiques ne peuvent pas différencier sans ambiguïté F. magna de « F. nericia »
Réactions biochimiques de la GPAC, y compris des souches de Finegoldia sp. ont été décrits; seulement peu de variations biochimiques chez Finegoldia sp. des souches ont déjà été signalées2,7. Nous avons testé deux systèmes de test biochimique disponibles dans le commerce qui sont utilisés pour l’identification des anaérobies. Le kit « API® 20A » contient 21 tests, dont 16 tests d’utilisation des glucides. Comme indiqué précédemment 7, aucun des glucides n’a pu être métabolisé par Finegoldia sp. déformation (données non représentées). Les autres réactions (indole, uréase, catalase, hydrolyse de l’esculine et liquéfaction de la gélatine) n’ont pas non plus révélé de différence entre les souches de F. magna et de « F. nericia » (données non montrées). Le kit « RapIDTM ANA II » contient 18 tests d’activités enzymatiques. Alors que les enzymes saccharolytiques font défaut, tous les Finegoldia sp. les souches produisent des enzymes protéolytiques (tableau S5). La réaction de détection de l’activité phosphatase était la seule réaction qui différait d’une souche à l’autre, toutes les souches de F. magna étant apparemment négatives et la plupart des souches de » F. nericia » positives. Cependant, l’interprétation de ce résultat de test est limitée car le changement de couleur était à la limite de la signification. En conclusion, deux systèmes de tests biochimiques commerciaux existants, API® 20 A et RapIDTM ANA II, ne peuvent pas différencier sans ambiguïté F. magna de « F. nericia ». Les propriétés biochimiques de Finegoldia sp. besoin d’être exploré plus avant avec des tests biochimiques supplémentaires.
La microscopie révèle des différences de surface entre F. magna et « F. nericia »
Pour tenter de visualiser la morphologie et la structure de surface de F. magna et « F. nericia » nous avons utilisé la microscopie électronique à balayage (MEB) et la microscopie à force atomique (AFM) sur des souches de F. magna (07T609, 08T492 et 09T408) et de « F. nericia » (09T494, 12T272 et 12T306). Le MEB a détecté des différences de surface : les cellules de F. magna étaient plus adhérentes ou agrégatives, produisant apparemment une matrice polymère extracellulaire (Fig. 6). Cela confirme des travaux antérieurs qui ont permis de visualiser une matrice d’exopolysaccharides dans des biofilms de F. magna (souche FmBs12) par microscopie électronique à balayage ainsi que par microscopie confocale à balayage laser23. Dans le test « F. nericia » souches nous n’avons pas pu identifier une telle matrice; les cellules semblent moins agrégatives.
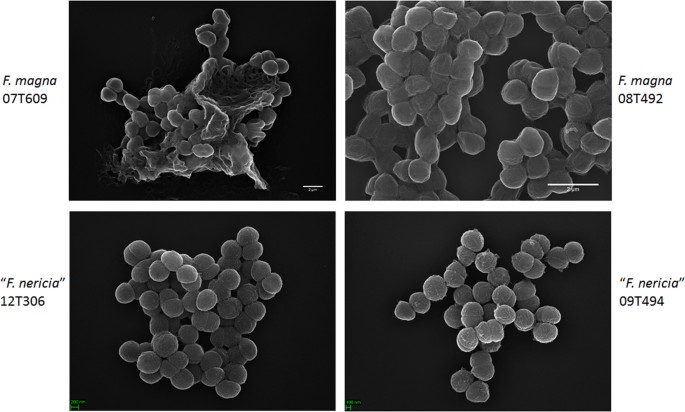
Comparaison au microscope électronique à balayage de F. magna et de « F. nericia ». Les panneaux supérieur et inférieur montrent des images représentatives de F. magna 07T609 et 08T492 et de « F. nericia » 09T494 et 12T306, respectivement. Les cellules des souches de F. magna sont plus adhérentes/agrégatives et semblent produire une matrice polymère, en particulier la souche 07T609; en revanche, les cellules de « F. nericia » sont moins agrégatives.
L’AFM a confirmé ces observations. De plus, nous avons observé des différences concernant les appendices cellulaires. Les cellules de F. magna produisent des appendices filamenteux plus longs qui dépassent de toute la surface cellulaire; ces structures pourraient représenter des pili ou des fimbriae (Fig. 7). En revanche, les cellules des souches de « F. nericia » avaient une surface plus lisse sans longs appendices saillants. Il n’est cependant pas exclu que ces observations AFM dépendent des conditions de culture bactérienne et des étapes de traitement des échantillons.

Comparaison microscopique à force atomique de F. magna et de « F. nericia ». Les panneaux de gauche et de droite représentent des images de F. magna 07T609 et de « F. nericia » 12T306, respectivement. (A et C) Images haute résolution avec méthode avancée assistée au toucher doux avec des barres d’échelle Z jusqu’à 0,3 µm et des barres XY indiquant 500 nm. (B et D) Superposition d’images d’images à contraste élevé de (A et C), combinées à une suppression polynomiale horizontale et verticale indépendante de (A et C), résolues au 11e degré. Des expériences ont été réalisées sur trois échantillons de chaque souche.