- genomen van stammen van Finegoldia
- op basis van genoomvergelijking
- Stamspecifieke heterogeniteit van Finegoldia sp
- Host-interagerende factoren en (vermeende) virulentiefactoren
- Finegoldia-genomen coderen functionele CAMP-factoren
- twee biochemische testsystemen kunnen F. magna niet ondubbelzinnig onderscheiden van”F. nericia”
- microscopie onthult oppervlakteverschillen tussen F. magna en”F. nericia”
genomen van stammen van Finegoldia
in totaal 17 genomen van Finegoldia sp. werden geanalyseerd en vergeleken. Tien stammen zijn onlangs gesequenced door onze groep15; ze werden geïsoleerd van patiënten met orthopedische gewrichtsimplantaat-geassocieerde infecties in Zweden. Zeven genomen werden eerder gesequenced door anderen, met inbegrip van het gesloten genoom van de typestam ATCC 2932814. De eigenschappen van het genoom van alle tot nu toe gesequenced stammen van Finegoldia worden samengevat in Tabel 1. De annotatie met behulp van Prokka ontdekt tussen 1570 en 1906 codage sequenties (CDS) per genoom, met een gemiddelde van 1760 CDS per stam.
op basis van genoomvergelijking
merkten we eerder een grote heterogeniteit op tussen genomen van isolaten van Finegoldia 15. Een kern-genoom afstemming van alle 17 genomen werd gedaan en de enige nucleotide polymorfismen (SNPs) in het kerngenoom werden geroepen. In totaal, werden 126.647 kern-genoom SNPs geà dentificeerd en gebruikt voor phylogenomic wederopbouw, onthullend dat de 17 genomen in twee verschillende clades kunnen worden gegroepeerd (Fig. 1). Een aanvullende analyse die het gemiddelde nucleotide identificeert (ANI) van het kerngenoom bepaalt toonde aan dat de twee clades een ANI van 90,7%, met een stam-specifieke Ani variatie van 90,2% aan 91,2% tentoonstellen. Een dergelijke lage ANI geeft aan dat de twee clades individuele soorten vertegenwoordigen16. Eén clade bevat de meeste eerder gesequenced en bestudeerde stammen, waaronder ATCC29328 en ALB8; het kan worden beschouwd als de klassieke F. magna clade, met een intraclade ANI van 96,6% (variatie 95,1–97,4%). De andere clade met een intraclade ANI van 94,8% (93,4–96.6%) bevat een andere ATCC-stam (ATCC 53516)en voornamelijk stammen geïsoleerd van patiënten in het Örebro Universitair Ziekenhuis, Zweden15. Hierna worden stammen van deze clade voorlopig “Finegoldia nericia” (Latijn voor Närke, de Zweedse regio, waar de meeste stammen van deze soort werden geïsoleerd) genoemd.

Fylogenomic boom op basis van kern-genoom SNPs van alle 17 sequenced genomen van Finegoldia stammen. Het programma Pastnp werd gebruikt om de kern genoom uit te lijnen en te bellen SNPs. Het kerngenoom wordt vertegenwoordigd door 43% van het referentiegenoom (ATCC29328); een totaal van 126.647 betrouwbare kern-genoom SNPs werden gebruikt om een geheel-genoomfylogenie te reconstrueren. Genomen kunnen worden toegewezen aan twee hoofdkladen; een clade vertegenwoordigt F. magna stammen en de andere clade bestaat uit stammen van een nieuwe soort, hier voorlopig genaamd “F. nericia”.
een vergelijking van 16S rRNA sequenties van alle isolaten toonde een hoge mate van algemene overeenkomst (>99%), maar detecteerde een scheiding van F. magna isolaten van “F. nericia” isolaten, als gevolg van 5 tot 9 “F. nericia ” – specifieke SNPs (figuur S1). Het bleek ook een grotere diversificatie onder “F. nericia” isolaten in vergelijking met F. magna. Vervolgens analyseerden we extra 16S rRNA-sequenties die behoren tot Finegoldia sp. opgeslagen in GenBank (figuur S2). Dit onthulde dat de meeste opgeslagen opeenvolgingen cluster met “F. nericia”, in het bijzonder een groep van Finegoldia sp. stammen geïsoleerd uit gal stent biofilms 17. Ook hier wordt een grotere diversificatie onder “F. nericia” isolaten gedetecteerd. Vervolgens analyseerden we menselijke microbioomgegevens opgeslagen in de IMG-database (Integrated Microbial genomen and microbioom samples) voor de aanwezigheid van F. magna en “F. nericia” sequenties. 3287 en 7068 coderende sequenties van F. magna ATCC29328 en” F. nericia ” ATCC 53516, respectievelijk, konden in de 875 geassembleerde menselijke microbioom datasets huidig in IMG worden geà dentificeerd (gegevens niet getoond); hoogste voorkomen van Finegoldia werden ontdekt in retroauriculaire vouw specimens. Bij elkaar genomen suggereren deze analyses dat bij de mens geassocieerde Finegoldia-isolaten “F. nericia” vaker voorkomt dan F. magna.
Stamspecifieke heterogeniteit van Finegoldia sp
om meer licht te werpen op de populatiestructuur van het genus Finegoldia werden vergelijkende analyses uitgevoerd van 12 genomen (alle tien nieuw gesequenced stammen en de twee ATCC-stammen), waaronder vier stammen van de F. magna clade en acht stammen van de “F. nericia” clade. Een visuele vergelijking van de 12 genomen benadrukte het nucleotide identiteits verschil tussen de twee clades (Fig. 2). Interessant is dat het plasmide gevonden in ATCC 29328 specifiek lijkt te zijn voor deze stam aangezien geen enkele andere sequenced stam een soortgelijke sequentie bevat (Fig. 2 bis).

genoom vergelijking van 12 stammen behorend tot het geslacht Finegoldia. De twee binnenste ringen vertegenwoordigen de G + C-inhoud (Zwart) en de GC-schuin (violet/groen). (A) de referentiestam is ATCC29328 (F. magna); let op de hoge nucleotide-identiteit (gevisualiseerd door de donkerrode kleur) van de drie binnenste genomen (die allen tot F. behoren. magna), en de onderste identificeren aan acht stammen behorend tot ” F. nericia “(lichtrode kleur van de acht buitenste ringen); van binnen naar buiten: stammen 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 en ATCC 53516. De grote ATCC29328 – specifieke regio in de linkerbovenhoek vertegenwoordigt de plasmide pFMC. B) de referentiestam is 09T494 (“F. nericia”). De zeven” F. nericia ” genomen (binnenringen) hebben een hoger nucleotide identificeren (vertegenwoordigd door de donkerdere rode kleur), in vergelijking met de vier F. magna genomen (buitenringen); van binnen naar buiten: stammen 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 en 09T408. Er zijn geen tekenen van “F. nericia”-specifieke genomische gebieden, maar let op de vijf grote stam-specifieke clusters.
er werden geen significante verschillen in genoomgrootte en aantal CDS ontdekt tussen de twee clades. Bovendien werden geen clade-specifieke (d.w.z. F. magna of “F. nericia”-specifieke) genomische regio ‘ s gedetecteerd, zoals beoordeeld uit de BRIG-analyse (Fig. 2) en ook bevestigd door een bidirectionele blast-benadering van alle CD ‘ s met Proteïnortho (tabel S1). Deze benadering ontdekte 1202 orthologs gedeeld door alle 12 genomen, d.w.z. in gemiddelde 68% van CDS van elke stam zijn deel van het kernproteoom. Er worden slechts zeer weinig clade-specifieke genen gevonden. In plaats daarvan kunnen veel regio ‘ s worden gevonden die slechts door een paar stammen worden gedeeld, bijvoorbeeld voor stam 09T494 (zie ook Fig. 2B). Daarnaast zijn stamspecifieke regio ‘ s aanwezig die in totaal 1016 stamspecifieke CDS coderen, met een groot bereik tussen de stammen, bijvoorbeeld 187 en 22 stamspecifieke CDS worden gedetecteerd in stam ATCC29328 en T151023, respectievelijk (tabel S1). Tot de stamspecifieke functies behoren diverse transportfuncties (ijzer, effluxpompen, oligopeptide ABC transporters), oppervlakte-eiwitten, restrictie-modificatiesystemen, conjugatieve overdrachtsfuncties, polyketidesynthese, bacteriocinen, faaggerelateerde functies, antibiotische resistentiedeterminanten en CRISPR/cas-systemen.
interessant is dat verschillen in de CRISPR/cas-systemen werden gedetecteerd tussen stammen van F. magna en “F. nericia” (tabel S2). Zeven van de negen F. magna stammen bevatten een tot vier CRISPR arrays per stam, met in gemiddelde 12 spacers (variërend van 3 tot 38 spacers per stam). De herhaling “GTTTGAGAATGATGTAATTTCATATAGGTATTAAC” was specifiek voor stammen van F. magna. In tegenstelling, “F. nericia” stammen droeg slechts één CRISPR array per stam die in gemiddelde 53 spacers (range 14 tot 87 per stam). Elke stam bevatte een individuele set van spacers in hun CRISPR array( s), onderliggende stam-specifieke heterogeniteit, en aangeeft dat elke stam heeft een unieke evolutionaire geschiedenis.
om te beoordelen of dergelijke stamspecifieke functies horizontaal werden verworven, hebben we met behulp van het IslandViewer-programma gekeken naar tekenen van hun mobiliteit/acquisitie (figuur S3). Dit programma voorspelde 141 (stam 09T408) tot 408 (stam 12T273) genen gelegen in genomische eilanden die waarschijnlijk horizontaal worden verworven, met inbegrip van veel van de bovengenoemde stam-specifieke genen betrokken bij specialisatie, fitness, overleving en interspecies competitie (tabel S3).
Host-interagerende factoren en (vermeende) virulentiefactoren
verschillende host-interagerende factoren zijn beschreven voor F. magna 4,5. Sortase-afhankelijke pili zijn geïdentificeerd die belangrijk zijn voor adhesie en kolonisatie13. Een genomische locus die codeert voor de genen voor de pilus subeenheid Fmp1, de vermeende subeenheid Fmp2 en drie sortasen wordt bewaard onder alle sequenced genomen (Fig. 3A). Er zijn strain-specifieke variaties up-en downstream van deze locus, d.w.z. sommige stammen bezitten extra genen stroomopwaarts van fmp1 en stroomafwaarts van de sortaseplaats, met inbegrip van een gen dat een hemolysin III-familieproteã ne codeert. Interessant is dat er strain-specific verschillen zijn met betrekking tot fmp1. Bijna elke stam draagt een individuele variant en een phylogenetic analyse van Fmp1 op eiwitniveau toonde geen correlatie met de phylogenomic analyse van het kerngenoom (Fig. 3B). Fmp2 daarentegen wordt bij alle stammen in hoge mate geconserveerd. Een ander eiwit, dat stroomopwaarts van fmp1 wordt gecodeerd, herbergt collageen – en fibrinogeenbindingsdomeinen; het collageen-bindend domein (COG4932) is ook aanwezig in Fmp2. Wij stellen voor dat dit eiwit ook tot de pilus locus behoort en het corresponderende gen fmp3 noemt.

Sortase-afhankelijke pili-codeert genomische loci in de genomen van Finegoldia stammen. (A) genomische organisatie van de loci in F. magna (rood) en “F. nericia” (Lichtblauw) stammen. De drie sortases genen zijn afgebeeld in lichtgroen (met het label “2”); het gen voor de belangrijkste pilus subeenheid Fmp1 is weergegeven in rood (met het label “1”) en fmp2 is afgebeeld in Oranje (met het label “3”). Stroomopwaarts van fmp1 en stroomafwaarts van de sortasegenen in sommige stammen zijn extra genen aanwezig (gelabeld “8”, hemolysine III-familieproteïne en “10”, hypothetisch eiwit). Een vermoedelijk collageenbindend eiwit, Fmp3 (gelabeld “4”), wordt vóór fmp1 gecodeerd. B) fylogenetische boom van Fmp1 van finegoldia stammen. De meeste stammen bezitten een individuele Fmp1 variant. De evolutionaire geschiedenis werd afgeleid door het gebruik van de maximale waarschijnlijkheid methode, gedaan in MEGA7.
eiwit L is waarschijnlijk de best bestudeerde factor van F. magna 8. Het is een superantigen dat B-cellen en immunoglobulinen richt. Het is ontdekt in ongeveer 10% van de tot nu toe geteste finegoldia isolaten, maar varieert in grootte, en aantallen van antigeen-bindende fragment (Fab) – bindende domeinen (B1 domein, pfam02246) 4,8,18. Een blast search met de bestudeerde proteïne L van Stam 312 (locus ID: m86697, 719 aa) onthult een paar zeer vergelijkbare (eiwitidentiteit boven 50%) proteïnen met verschillende groottes (603 tot 1417 aa) in een paar stammen, d.w.z. stammen CCUG54800, T161024, ATCC53516, 12T273 en 12T306 (Fig. 4A). Dit zijn allemaal” F. nericia ” stammen, wat erop wijst dat de B-cel superantigen vaker voorkomt bij deze soort. De uitzonderlijk grote (1417 aa) eiwit-l-achtige factor in stam 12T306 bezit naast twee B1-domeinen ook twee ga-modules die bekend zijn van peptostreptokokken albumine-bindende eiwitten (zie hieronder). Veel extra proteã nen met lage gelijkenis aan proteã ne L, maar zonder enige schijnbare B1 domeinen te bevatten worden gecodeerd in de genoom-sequenced finegoldia stammen (tabel S4A); nogmaals, stam-specifieke variaties bestaan, en een paar stammen, dat wil zeggen 09T408, 09T494 en T151023 bezitten geen dergelijke homologen.

genomische loci coderen eiwit L en FAF homologen in Finegoldia stammen. (A) Blast searching met het gekarakteriseerde eiwit L van Stam 312 (locus ID: M86697) werd uitgevoerd in genoom-sequenced finegoldia stammen, en homologen (in rood, gelabeld “1”) met een hoge gelijkenis (>50% eiwitidentiteit) worden getoond (zie ook Tabel S4A). Alleen “F. nericia” stammen, maar niet F. magna stammen bevatten dergelijke homologen met verschillende lengtes. B) Blast-zoekopdrachten met het bestudeerde FAF-eiwit van Stam ALB8 vonden homologen van volledige lengte (in rood, met het label “1”) in drie F. magna (rood) en vier “F. nericia” (Lichtblauw) stammen. Het gen FAF is frameshifted in de twee” F. nericia ” stammen T151023 en 12T272.
Finegoldia magna werd gevonden om aan menselijk albumin9 te binden. Een peptostreptokokken albumine-bindend eiwit (PAB) is onderzocht in de stam ALB8; de activiteit ervan is gekoppeld aan een versnelde groeisnelheid 9. Het PAB-eiwit van Stam ALB8 (locus ID: X77864) bevat GA-modules, het albumine-bindend domein; de structuur van de ga-module is resolved19. Een Blast search bleek dat een nauwe PAB homolog niet kan worden gevonden in andere sequenced Finegoldia genomen (tabel S4B). In plaats daarvan zijn er verschillende homologen die tot 50% eiwitidentiteit delen; de gelijkenissen van homologen in F. magna stammen zijn gemiddeld hoger vergeleken met homologen in “F. nericia” stammen. Met betrekking tot ga-module-bevattende eiwitten, worden in totaal 16 eiwitten gecodeerd in de 12 Finegoldia genomen, wat wijst op enige redundantie van albumine-bindende eiwitten (tabel S4C). De meeste stammen coderen twee eiwitten met GA-modules, behalve van Stam 09T408 (F. magna) en t151023 (“F. nericia”).
het celwandgebonden subtilisine-achtige protease SufA is aangetoond dat het verschillende menselijke eiwitten afbreekt, zoals collageen IV, fibrinogeen en antimicrobiële peptiden zoals LL-37 en MIG/CXCL910,20,21. Blast-zoekopdrachten stelden vast dat de meeste Finegoldia-stammen een Sufa-homolog hebben die sterk lijkt (eiwitidentiteit > 75%) op de onderzochte SufA van Stam ALB8 (locus ID: DQ679960). Uitzonderingen zijn de stammen 07T609, 09T408 en CCUG54800. Een tweede SufA homolog is aanwezig in de meeste stammen; het heeft een gemiddelde eiwitidentiteit van 26% (tabel S4D).
een andere belangrijke host-interagerende factor is FAF (F. magna adhesiefactor). Rond 90% van F. magna stammen produceren deze factor, die verantwoordelijk is voor het samenklonteren van bacteriën en bemiddelt band aan het kelderverdiepingsmembraan door aan BM-4011 te binden. We vonden dat zeven (drie F. magna en vier “F. nericia” stammen) van de 12 stammen coderen voor een FAF homolog vergelijkbaar met die bestudeerd in de ALB8 stam (Fig. 4B). Stam-specifieke verschillen bestaan en frameshiftmutaties worden gezien in” F. nericia ” stammen T151023 en 12T272. Er is geen homolog gevonden in de stammen 09T408, 09T494 en 12T306 (tabel S4E).
Finegoldia-genomen coderen functionele CAMP-factoren
we zochten in het genoom naar andere veronderstelde gastheer-interacterende factoren en vonden genen die coderen voor Christie-Atkins-Munch-Petersen (CAMP) – factoren. CAMP factoren kunnen fungeren als co-hemolysines en verantwoordelijk zijn voor de zogenaamde CAMP reactie, de synergetische lysis van schapen erytrocyten door Staphylococcus aureus sphingomyelinase C (beta-toxine) en een CAMP factor22.
Genoomwinning identificeerde twee genen die coderen voor CAMP-factoren in elke Genoomgesequenced finegoldia-stam (CAMP1 en CAMP2; figuur S4); elke homolog herbergt het CAMP-factor-familiedomein (Pfam07373). Interessant is dat één homolog (CAMP2) per stam een extra bacterieel IG-achtig domein herbergt (Pfam02368). Naast CAMP1 en CAMP2, vier “F. nericia ” stammen bevatten twee extra kamp factoren die wezenlijk verschillen van de andere homologen (CAMP3 en CAMP4; figuur S4). Dus vier van de zeven “F nericia” stammen bevatten vier CAMP factor genen.
vervolgens wilden we weten of Finegoldia-stammen een positieve CAMP-reactie veroorzaken. Een test op de CAMP agarplaat toont aan dat sommige stammen sterk CAMP-reactiepositief zijn, terwijl voor andere stammen slechts weinig co-hemolyse kan worden gedetecteerd onder de toegepaste kweekomstandigheden na twee en vijf dagen incubatie (Fig. 5, Figuur S5). Alle drie getest F. magna stammen waren sterk kamp reactie-positief. Onder “F. nericia” stammen ontdekten we enige heterogeniteit; twee stammen waren sterk CAMP reactie-positief, vier stammen waren slechts licht co-hemolytische, en een stam, CCUG54800, was duidelijk CAMP reactie-negatief. De Genomic analyse vond dat in stam CCUG54800 het CAMP1-codeert gen een toevoeging van een transposase-codeert locus in het 5′-eind van het gen bevatte, dat het gen verstoort (figuur S6).

CAMP reactie van Finegoldia stammen. TSA agar platen met 5% schapenbloed werden gebruikt. Staphylococcus aureus wordt geïnoculeerd in de middelste streep. Een positieve CAMP-test wordt aangetoond door volledige erytrocytenlysis op het raakvlak van de Finegoldia sp. en de S. aureus strepen. Alle drie de F. magna (in rood) stammen en twee van de zeven “F. nericia” (in blauw) stammen vertoonden een sterke positieve CAMP reactie na twee dagen anaërobe incubatie.
onze resultaten geven aan dat CAMP factoren van Finegoldia sp. kan functioneel zijn en dat er stam-specifieke verschillen in CAMP factor activiteit.
twee biochemische testsystemen kunnen F. magna niet ondubbelzinnig onderscheiden van”F. nericia”
biochemische reacties van GPAC inclusief stammen van Finegoldia sp. zijn beschreven; slechts weinig biochemische variatie onder Finegoldia sp. stammen werden eerder vermeld2, 7. We hebben twee commercieel verkrijgbare biochemische testsystemen getest die worden gebruikt voor de identificatie van anaëroben. De kit “API ® 20A” bevat 21 tests, waaronder 16 tests voor het gebruik van koolhydraten. Zoals eerder gemeld 7, kon geen van de koolhydraten worden gemetaboliseerd door Finegoldia sp. stam (gegevens niet weergegeven). De andere reacties (indool, urease, catalase, esculine hydrolyse en gelatine liquefactie) lieten ook geen verschil zien tussen stammen van F. magna en “F. nericia” (gegevens niet getoond). De kit “RapIDTM ANA II” bevat 18 testen voor enzymatische activiteiten. Terwijl saccharolytische enzymen ontbreken, zijn alle Finegoldia sp. stammen produceren proteolytische enzymen (tabel S5). De reactie die fosfatase-activiteit detecteerde was de enige reactie die verschilde tussen de stammen, met alle F. magna-stammen blijkbaar negatief en de meeste “F. nericia” – stammen positief. De interpretatie van dit testresultaat is echter beperkt omdat de kleurverandering zich op de grens van betekenis bevond. Kortom, twee bestaande commerciële biochemische testsystemen, API ® 20 A en RapIDTM ANA II, kunnen F. magna niet ondubbelzinnig onderscheiden van”F. nericia”. De biochemische eigenschappen van Finegoldia sp. moeten verder worden onderzocht met aanvullende biochemische tests.
microscopie onthult oppervlakteverschillen tussen F. magna en”F. nericia”
In een poging om de morfologie en oppervlaktestructuur van F. magna en ” F. nericia ” we gebruikten scanning elektronenmicroscopie (SEM) en atomic force microscopy (AFM) op stammen van F. magna (07T609, 08T492, en 09T408) en “F. nericia” (09T494, 12T272 en 12T306). SEM ontdekte oppervlakteverschillen: cellen van F. magna waren meer adherent of aggregatief, schijnbaar producerend een extracellulaire polymeermatrix (Fig. 6). Dit bevestigt vorig werk dat een exopolysaccharidematrijs in biofilms van F. magna (stam FmBs12) door het aftastenelektronenmicroscopie evenals confocal laserscanning microscopie23 heeft gevisualiseerd. In de geteste “F. nericia “stammen we konden zo’ n matrix niet identificeren; cellen lijken minder aggregatief.
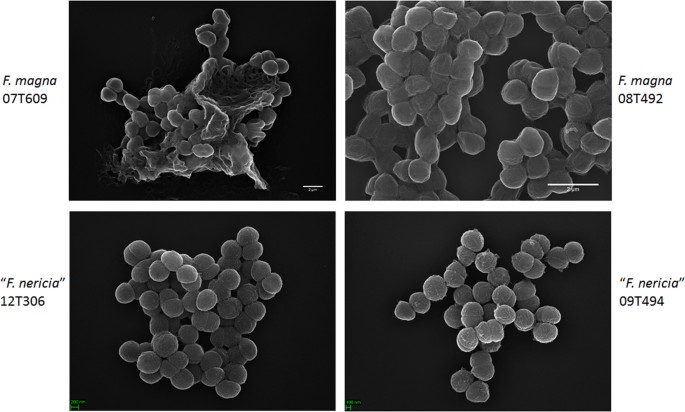
Scanning electron microscopic comparison of F. magna and “F. nericia”. De bovenste en onderste panelen tonen representatieve beelden van F. magna 07T609 en 08T492 en” F. nericia ” 09T494 en 12T306, respectievelijk. Cellen van F. magna stammen zijn meer adherent / aggregatief en lijken een polymeermatrix te produceren, in het bijzonder stam 07T609; daarentegen zijn cellen van “F. nericia” minder aggregatief.
de AFM heeft deze waarnemingen bevestigd. Bovendien zagen we verschillen met betrekking tot celaanhangsels. Cellen van F. magna produceerden langere filamenteuze aanhangsels die uit het gehele celoppervlak uitsteekten; deze structuren zouden pili of fimbriae kunnen vertegenwoordigen (Fig. 7). In tegenstelling, cellen van “F. nericia” stammen hadden een gladder oppervlak zonder lange uitstekende aanhangsels. Het valt echter niet uit te sluiten dat deze waarnemingen van de AFM afhankelijk zijn van de bacteriële kweekomstandigheden en de monsterverwerkingsstappen.

Atomic force microscopic comparison of F. magna and “F. nericia”. De linker en rechter panelen vertegenwoordigen beelden van F. magna 07T609 en “F. nericia” 12T306, respectievelijk. (A en C) beelden met hoge resolutie met geavanceerde soft-touch-methode met z-balken tot 0,3 µm en XY-balken die 500 nm aangeven. (B en D) Overlay beelden van hoog contrast beelden van (A en C), gecombineerd met onafhankelijke horizontale en verticale polynoom verwijdering van (A en C), opgelost tot de 11e graad. Experimenten werden uitgevoerd op drie monsters van elke stam.