- genomer af stammer af Finegoldia
- opdeling af Finegoldia stammer i to forskellige klader baseret på genom sammenligning
- Stammespecifik heterogenitet af Finegoldia sp
- Host-interagerende faktorer og (formodede) virulensfaktorer
- Finegoldia-genomer koder for CAMP-faktorer, der er funktionelle
- to biokemiske testsystemer kan ikke entydigt differentiere F. magna fra “F. nericia”
- mikroskopi afslører overfladeforskelle mellem F. magna og”F. nericia “
genomer af stammer af Finegoldia
i alt 17 genomer af Finegoldia sp. blev analyseret og sammenlignet. Ti stammer er for nylig blevet sekventeret af vores gruppe15; de blev isoleret fra patienter med ortopædiske ledimplantatassocierede infektioner i Sverige. Syv genomer blev tidligere sekventeret af andre, herunder det lukkede genom af typestammen ATCC 2932814. Genomegenskaber af alle hidtil sekventerede stammer af Finegoldia er opsummeret i tabel 1. Annotationen ved hjælp af Prokka detekterede mellem 1570 og 1906 kodende sekvenser (CDS) pr.genom med et gennemsnit på 1760 CDS pr. stamme.
opdeling af Finegoldia stammer i to forskellige klader baseret på genom sammenligning
vi har tidligere bemærket en stor heterogenitet blandt genomer af Finegoldia isolater15. En kerne-genomjustering af alle 17 genomer blev udført, og enkelt nukleotidpolymorfier (SNP ‘ er) i kernegenomet blev kaldt. I alt blev 126.647 kerne-genom SNP ‘ er identificeret og anvendt til fylogenomisk rekonstruktion, hvilket afslørede, at de 17 genomer kan grupperes i to forskellige klader (Fig. 1). En yderligere analyse, der bestemmer den gennemsnitlige nukleotididentifikation (ANI) af kernegenomet, viste, at de to klader udviser en ANI på 90,7% med en stammespecifik ANI-variation fra 90,2% til 91,2%. En sådan lav ANI indikerer, at de to klader repræsenterer individuelle arter16. En clade indeholder de fleste tidligere sekventerede og studerede stammer, herunder atcc29328 og ALB8; det kan betragtes som den klassiske F. magna clade, med en intraclade ANI på 96,6% (variation 95,1–97,4%). Den anden klade med en intraclade ANI på 94,8% (93,4–96.6%) indeholder en anden ATCC-stamme (ATCC 53516) og hovedsageligt stammer isoleret fra patienter på Sveriges Universitetshospital, Sverige15. Herefter benævnes stammer af denne klade foreløbigt “Finegoldia nericia” (Latin for N Larrke, den svenske region, hvor de fleste stammer af denne art blev isoleret).

Fylogenomisk træ baseret på kerne-genom SNP ‘ er af alle 17 sekventerede genomer af Finegoldia-stammer. Programmet Pastinp blev brugt til at justere kernegenomet og kalde SNP ‘ er. Kernegenomet er repræsenteret af 43% af referencegenomet (atcc29328); i alt 126.647 pålidelige kernegenom SNP ‘ er blev brugt til at rekonstruere en helgenom fylogeni. Genomer kan tildeles to hovedklader; den ene klade repræsenterer F. magna-stammer, og den anden klade består af stammer af en ny art, her foreløbigt navngivet “F. nericia”.
en sammenligning af 16S rRNA-sekvenser af alle isolater afslørede en høj grad af samlet lighed (>99%), men detekterede en adskillelse af F. magna-isolater fra “F. nericia” – isolater på grund af 5 til 9 “F. nericia “- specifikke SNP ‘ er (Figur S1). Det afslørede også en højere diversificering blandt “F. nericia” isolater sammenlignet med F. magna. Dernæst analyserede vi yderligere 16S rRNA-sekvenser tilhørende Finegoldia sp. gemt i GenBank (figur S2). Dette afslørede, at de fleste lagrede sekvenser klynge med “F. nericia”, især en gruppe af Finegoldia sp. stammer isoleret fra biofilm med galdestent17. Igen detekteres en højere diversificering blandt” F. nericia ” isolater. Dernæst analyserede vi humane mikrobiomdata gemt i IMG-databasen (Integrated Microbial Genomes and microbiome samples) for tilstedeværelsen af F. magna og “F. nericia” – sekvenser. 3287 og 7068 kodende sekvenser af henholdsvis F. magna atcc29328 og” F. nericia ” ATCC 53516 kunne identificeres i de 875 samlede humane mikrobiomdatasæt, der er til stede i IMG (data ikke vist); højeste forekomster af Finegoldia blev påvist i retroaurikulære foldeprøver. Samlet set antyder disse analyser, at blandt humanassocierede Finegoldia isolater” F. nericia ” synes at være mere udbredt end F. magna.
Stammespecifik heterogenitet af Finegoldia sp
for at kaste yderligere lys over populationsstrukturen i Finegoldia-slægten blev der udført sammenlignende analyser af 12 genomer (alle ti nyligt sekventerede stammer og de to ATCC-stammer), herunder fire stammer af F. magna clade og otte stammer af “F. nericia” clade. En visuel sammenligning af de 12 genomer fremhævede nukleotididentitetsforskellen mellem de to klader (Fig. 2). Interessant nok synes plasmidet, der findes i ATCC 29328, at være specifikt for denne stamme, da ingen anden sekventeret stamme indeholder en lignende sekvens (Fig. 2A).

genom sammenligning af 12 stammer tilhørende slægten Finegoldia. De to inderste ringe repræsenterer G + C-indholdet (sort) og GC-skævheden (violet/grøn). (A) referencestammen er ATCC29328 (F. magna); bemærk den høje nukleotididentitet (visualiseret af den mørkerøde farve) af de tre inderste genomer (der tilhører alle F. til otte stammer, der tilhører “F. nericia” (lys rød farve på de otte ydre ringe); indefra og udefra: stammer 07T609, 08t492, 09t408, 09t494, 12t272, 12t273, 12t306, CCUG 54800, T151023, T160124 og ATCC 53516. Den store atcc29328-specifikke region i den øverste venstre region repræsenterer plasmidet pFMC. B) referencestammen er 09T494 (“F. nericia”). De syv” F. nericia ” – genomer (indre ringe) har en højere nukleotididentifikation (repræsenteret af den mørkere røde farve) sammenlignet med de fire F. magna-genomer (ydre ringe); indefra og udefra: stammer 12T272, ATCC 53516, 12t306, 12t273, CCUG 54800, T151023, T160124, atcc29328, 07t609, 08t492 og 09t408. Der er ingen tegn på “F. nericia”-specifikke genomiske regioner, men bemærk de fem store stammespecifikke klynger.
der blev ikke påvist nogen signifikante forskelle i genomstørrelse og antal CDS mellem de to klader. Derudover blev der ikke påvist nogen clade-specifik (dvs.F. magna eller “F. nericia”-specifik) genomiske regioner, som bedømt ud fra BRIG-analysen (Fig. 2) og også bekræftet af en tovejs blast tilgang af alle cd ‘ er ved hjælp af ProteinOrtho (tabel S1). Denne fremgangsmåde detekterede 1202 ortologer, der deles af alle 12 genomer, dvs.i gennemsnit er 68% af CDS ‘ erne for hver Stamme en del af kerneproteomet. Kun meget få clade-specifikke gener findes. I stedet findes mange regioner, der kun deles af nogle få stammer, f.eks. for Stamme 09T494 (se også Fig. 2B). Derudover er der stammespecifikke regioner til stede, der koder i alt 1016 stammespecifikke cd ‘er med et stort interval mellem stammerne, f.eks. 187 og 22 stammespecifikke cd’ er detekteres i henholdsvis stamme atcc29328 og T151023 (tabel S1). Blandt de stammespecifikke funktioner er forskellige transportfunktioner (jern, efflukspumper, oligopeptid ABC-transportører), overfladeproteiner, restriktionsmodifikationssystemer, konjugative overførselsfunktioner, polyketidsyntese, bakteriociner, fagrelaterede funktioner, antibiotikaresistensdeterminanter og CRISPR/cas-systemer.
interessant nok blev forskelle i CRISPR/cas-systemerne påvist mellem stammer af F. magna og “F. nericia” (tabel S2). Syv ud af ni F. magna-stammer indeholdt en til fire CRISPR-arrays pr.stamme, der i gennemsnit indeholdt 12 afstandsstykker (fra 3 til 38 afstandsstykker pr. stamme). Gentagelsen” GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC ” var specifik for stammer af F. magna. I modsætning hertil bar “F. nericia” – stammer kun et CRISPR-array pr.stamme, der indeholdt i gennemsnit 53 afstandsstykker (interval 14 til 87 pr. stamme). Hver stamme indeholdt et individuelt sæt afstandsstykker i deres CRISPR-array(er), hvilket understregede stammespecifik heterogenitet og indikerer, at hver stamme har en unik evolutionær historie.
for at vurdere, om sådanne belastningsspecifikke funktioner blev erhvervet horisontalt, kiggede vi på tegn på deres mobilitet/erhvervelse ved hjælp af Islandviserprogrammet (figur S3). Dette program forudsagde 141 (stamme 09T408) til 408 (stamme 12t273) gener placeret på genomiske øer, der sandsynligvis vil blive erhvervet vandret, herunder mange af de ovennævnte stammespecifikke gener involveret i specialisering, fitness, overlevelse og interspecies konkurrence (tabel S3).
Host-interagerende faktorer og (formodede) virulensfaktorer
flere host-interagerende faktorer er blevet beskrevet for F. magna 4,5. Der er identificeret sortaseafhængige pili, som er vigtige for adhæsion og kolonisering13. Et genomisk locus, der koder for generne for pilus-underenheden Fmp1, den formodede underenhed Fmp2 og tre sortaser bevares blandt alle sekventerede genomer (Fig. 3A). Der er stammespecifikke variationer op-og nedstrøms for dette sted, dvs. nogle stammer besidder yderligere gener opstrøms for fmp1 og nedstrøms for sortase locus, inklusive et gen, der koder for et hæmolysin III-familieprotein. Interessant nok er der belastningsspecifikke forskelle med hensyn til fmp1. Næsten hver stamme bærer en individuel variant, og en fylogenetisk analyse af Fmp1 på proteinniveau viste ingen sammenhæng med den fylogenomiske analyse af kernegenomet (Fig. 3B). I modsætning hertil er Fmp2 stærkt bevaret blandt alle stammer. Et andet protein, kodet opstrøms for fmp1, har kollagen-og fibrinogenbindingsdomæner; det kollagenbindende domæne (COG4932) er også til stede i fmp2. Vi foreslår, at dette protein også hører til pilus locus og navngiver det tilsvarende gen fmp3.

Sortaseafhængig pili-kodende genomisk loci i genomerne af Finegoldia-stammer. (A) genomisk organisering af loci i F. magna (rød) og “F. nericia” (Lyseblå) stammer. De tre sortasegener er afbildet i lysegrøn (mærket “2”); genet for den store pilus-underenhed Fmp1 er vist i rødt (mærket “1”) og fmp2 er afbildet i orange (mærket “3”). Opstrøms for fmp1 og nedstrøms for sortase-generne i nogle stammer er yderligere gener (mærket “8”, hæmolysin III-familieprotein og “10”, hypotetisk protein) til stede. Et formodet kollagenbindende protein, Fmp3 (mærket “4”), kodes opstrøms for fmp1. (B) fylogenetisk træ af Fmp1 af Finegoldia stammer. De fleste stammer har en individuel fmp1-variant. Den evolutionære historie blev udledt ved hjælp af den maksimale Sandsynlighedsmetode, udført i MEGA7.
Protein L er nok den bedst studerede faktor af F. magna 8. Det er en superantigen, der er målrettet mod B-celler og immunoglobuliner. 10% af de hidtil testede Finegoldia-isolater, men varierer i størrelse og antal antigenbindende fragment (Fab)-bindende domæner (B1-domæne, pfam02246)4,8,18. En blast-søgning med det undersøgte protein L fra stamme 312 (locus ID: M86697, 719 aa) afslører et par meget ens (proteinidentitet over 50%) proteiner med forskellige størrelser (603 til 1417 aa) i nogle få stammer, dvs. stammer CCUG54800, T161024, ATCC53516, 12t273 og 12t306 (Fig. 4A). Alle disse er” F. nericia ” stammer, hvilket indikerer, at B-celle superantigen er mere udbredt i denne art. Den usædvanligt store (1417 aa) protein L-lignende faktor i stamme 12t306 besidder udover to B1-domæner også to GA-moduler, der er kendt fra peptostreptokokalbumbindende proteiner (se nedenfor). Mange yderligere proteiner med lav lighed med protein L, men uden at indeholde nogen tilsyneladende B1-domæner er kodet i de genomsekventerede Finegoldia-stammer (tabel S4A); igen findes der stammespecifikke variationer, og nogle få stammer, dvs.09T408, 09t494 og T151023, har ingen sådanne homologer.

genomiske loci koder for protein L og FAF homologer i Finegoldia stammer. (A) Blast-søgninger med det karakteriserede protein L fra stamme 312 (locus ID: M86697) blev udført i Genomsekventerede Finegoldia-stammer, og homologer (i rødt, mærket “1”) med høj lighed (>50% proteinidentitet) er vist (Se også tabel S4A). Kun” F. nericia ” stammer, men ikke F. magna-stammer indeholder sådanne homologer med varierende længder. (B) Blast-søgninger med det undersøgte FAF-protein af stamme ALB8 fandt homologer i fuld længde (i rødt, mærket “1”) i tre F. magna (rød) og fire “F. nericia” (Lyseblå) stammer. FAF genet er frameshifted i de to” F. nericia ” stammer T151023 og 12t272.
Finegoldia magna blev fundet at binde til humant albumin9. Et peptostreptokokalbuminbindende protein (PAB) er blevet undersøgt i stammen ALB8; dets aktivitet er knyttet til en accelereret væksthastighed9. PAB-proteinet af stammen ALB8 (locus ID: 77864) indeholder ga-moduler, det albuminbindende domæne; strukturen af GA-modulet er opløset19. En Blast-søgning afslørede, at en tæt PAB-homolog ikke kan findes i andre sekventerede Finegoldia-genomer (tabel S4B). I stedet er der flere homologer, der deler op til 50% proteinidentitet; ligheder mellem homologer i F. magna-stammer er i gennemsnit højere sammenlignet med homologer i “F. nericia” – stammer. Med hensyn til GA-modulholdige proteiner kodes i alt 16 proteiner i de 12 Finegoldia-genomer, hvilket indikerer en vis redundans af albuminbindende proteiner (tabel S4C). De fleste stammer koder for to proteiner med GA-moduler, undtagen fra stamme 09T408 (F. magna) og T151023 (“F. nericia”).
den cellevægbundne subtilisin-lignende protease SufA har vist sig at nedbryde flere humane proteiner, såsom kollagen IV, fibrinogen og antimikrobielle peptider,såsom LL-37 og mig/CHCL910,20, 21. Blast-søgninger fastslog, at de fleste Finegoldia-stammer bærer en SufA-homolog, der er meget ens (proteinidentitet > 75%) til den studerede SufA fra stamme ALB8 (locus ID: DK679960). Undtagelser er stammerne 07T609, 09t408 og CCUG54800. En anden SufA homolog er til stede i de fleste stammer; den har en gennemsnitlig proteinidentitet på 26% (tabel S4D).
en anden vigtig værtsinteragerende faktor er FAF (F. magna adhæsionsfaktor). Cirka 90% af F. magna-stammer producerer denne faktor, som er ansvarlig for klumpning af bakterier og formidler binding til kældermembranen ved binding til BM-4011. Vi fandt ud af, at syv (tre F. magna og fire “F. nericia” – stammer) ud af de 12 stammer koder for en FAF-homolog svarende til den, der blev undersøgt i ALB8-stammen (Fig. 4B). Der findes stammespecifikke forskelle, og frameshift-mutationer ses i “F. nericia” – stammerne T151023 og 12t272. Der findes ingen homolog i stammerne 09T408, 09t494 og 12t306 (tabel S4E).
Finegoldia-genomer koder for CAMP-faktorer, der er funktionelle
vi søgte genomet efter andre formodede værtsinteragerende faktorer og fandt gener, der koder for Christie-Atkins-Munch-Petersen (CAMP) faktorer. CAMP-faktorer kan fungere som co-hæmolysiner og redegøre for den såkaldte CAMP-reaktion, den synergistiske lysis af fårerytrocytter ved Staphylococcus aureus sphingomyelinase C (beta-toksin) og en CAMP-faktor22.
Genomminedrift identificerede to gener, der koder for CAMP-faktorer i hver Genomsekventeret Finegoldia-stamme (CAMP1 og CAMP2; figur S4); hver homolog har CAMP factor-familiedomænet (Pfam07373). Interessant nok har en homolog (CAMP2) pr.stamme et yderligere bakterielt ig-lignende domæne (Pfam02368). Ud over CAMP1 og CAMP2, fire ” F. nericia ” – stammer indeholder to yderligere CAMP-faktorer, der adskiller sig væsentligt fra de andre homologer (CAMP3 og CAMP4; figur S4). Således har fire ud af syv “F. nericia” – stammer fire CAMPFAKTORGENER.
dernæst ønskede vi at vide, om Finegoldia-stammer producerer en positiv CAMPREAKTION. En CAMP agar plate assay viser, at nogle stammer er stærkt CAMP reaktionspositive, mens for andre stammer kun lidt Co-hæmolyse kan påvises under de anvendte dyrkningsbetingelser ved to og fem dages inkubation (Fig. 5, Figur S5). Alle tre testede F. magna-stammer var stærkt CAMP-reaktionspositive. Blandt” F. nericia ” – stammer opdagede vi en vis heterogenitet; to stammer var stærkt CAMP-reaktionspositive, fire stammer var kun lidt Co-hæmolytiske, og en stamme, CCUG54800, var klart CAMP-reaktionsnegativ. Genomisk analyse viste, at CAMP1-kodende gen i stamme CCUG54800 indeholdt en indsættelse af et transposase-kodende locus i 5′-enden af genet, som forstyrrer genet (figur S6).

CAMP reaktion af Finegoldia stammer. TSA-agarplader med 5% fåreblod blev anvendt. Staphylococcus aureus inokuleres i den midterste stribe. En positiv CAMP-test er indikeret ved fuldstændig erytrocytlysis ved grænsefladen til Finegoldia sp. og S. aureus-striberne. Alle tre F. magna (i røde) stammer og to ud af syv “F. nericia” (i blå) stammer viste en stærk positiv CAMPREAKTION efter to dages anaerob inkubation.
vores resultater indikerer, at CAMPFAKTORER for Finegoldia sp. kan være funktionel, og at der er belastningsspecifikke forskelle i CAMPFAKTORAKTIVITET.
to biokemiske testsystemer kan ikke entydigt differentiere F. magna fra “F. nericia”
biokemiske reaktioner af GPAC inklusive stammer af Finegoldia sp. er blevet beskrevet; kun lidt biokemisk variation blandt Finegoldia sp. stammer blev tidligere rapporteret2, 7. Vi testede to kommercielt tilgængelige biokemiske testsystemer, der bruges til identifikation af anaerober. Kittet” API Kurt 20A ” indeholder 21 tests, blandt dem 16 tests for kulhydratudnyttelse. Som rapporteret tidligere7 kunne ingen af kulhydraterne metaboliseres af nogen Finegoldia sp. stamme (data ikke vist). De andre reaktioner (indol, urease, catalase, esculinhydrolyse og gelatinefiktion) afslørede heller ikke nogen forskel mellem stammer af F. magna og “F. nericia” (data ikke vist). Kittet” RapIDTM ANA II ” indeholder 18 test for fermatiske aktiviteter. Mens saccharolytiske symptomer mangler, alle Finegoldia sp. stammer producerer proteolytiske stoffer (tabel S5). Reaktionen, der detekterede phosphataseaktivitet, var den eneste reaktion, der adskilte sig mellem stammerne, med alle F. magna-stammer tilsyneladende negative og de fleste “F. nericia” – stammer positive. Fortolkningen af dette testresultat er imidlertid begrænset, da farveændringen var på grænsen til Betydning. Afslutningsvis kan to eksisterende kommercielle biokemiske testsystemer, API Kart 20 A og RapIDTM Ana II, ikke entydigt differentiere F. magna fra “F. nericia”. De biokemiske egenskaber af Finegoldia sp. behov for at blive undersøgt yderligere med yderligere biokemiske tests.
mikroskopi afslører overfladeforskelle mellem F. magna og”F. nericia “
i et forsøg på at visualisere morfologien og overfladestrukturen af F. magna og ” F. nericia ” vi brugte scanningselektronmikroskopi (SEM) og atomkraftmikroskopi (AFM) på stammer af F. magna (07t609, 08t492 og 09T408) og “F. nericia” (09t494, 12t272 og 12t306). Sem detekterede overfladeforskelle: celler af F. magna var mere klæbende eller aggregerende, tilsyneladende producerer en ekstracellulær polymermatrice (Fig. 6). Dette bekræfter tidligere arbejde, der har visualiseret en eksopolysaccharidmatrice i biofilm af F. magna (stamme FmBs12) ved at scanne elektronmikroskopi såvel som konfokal laserscanningsmikroskopi23. I den testede ” F. nericia ” stammer vi kunne ikke identificere en sådan matrice; celler synes mindre aggregerende.
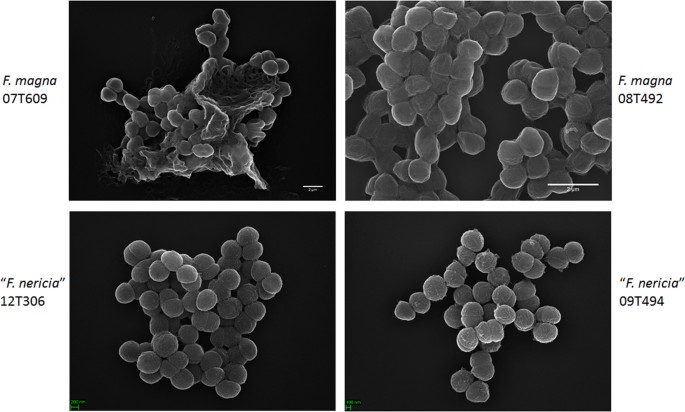
Scanning elektronmikroskopisk sammenligning af F. magna og”F. nericia”. De øverste og nederste paneler viser repræsentative billeder af henholdsvis F. magna 07t609 og 08t492 og “F. nericia” 09t494 og 12t306. Celler af F. magna-stammer er mere klæbende/aggregerende og synes at producere en polymermatrice, især stamme 07T609; i modsætning hertil er celler af “F. nericia” mindre aggregerende.
AFM bekræftede disse observationer. Desuden observerede vi forskelle med hensyn til cellevedhæng. Celler af F. magna producerede længere filamentøse vedhæng, der stak ud fra hele celleoverfladen; disse strukturer kunne repræsentere pili eller fimbriae (Fig. 7). I modsætning hertil havde celler af “F. nericia” – stammer en glattere overflade uden lange fremspringende vedhæng. Det kan imidlertid ikke udelukkes, at disse AFM-observationer afhænger af bakteriedyrkningsbetingelserne og af prøvebehandlingstrinnene.

atomic force mikroskopisk sammenligning af F. magna og”F. nericia”. De venstre og højre paneler repræsenterer billeder af henholdsvis F. magna 07t609 og “F. nericia” 12t306. (A og C) billeder i høj opløsning med assisteret avanceret soft-touch-metode med vægtstænger på op til 0,3 liter og vægtstænger, der angiver 500 nm. (B og D) Overlay billeder af høj kontrast billeder af (A og C), kombineret med uafhængig vandret og lodret polynomial fjernelse af (A og C), løst til 11.grad. Eksperimenter blev udført på tre prøver fra hver stamme.