총 17 의 유전자 Finegoldia sp. 분석 및 비교 하였다. 10 균주는 최근 우리 그룹에 의해 염기 서열되었습니다 15;그들은 스웨덴에서 정형 외과 관절 임플란트 관련 감염 환자에서 격리되었다. 7 개의 게놈은 이전에 다른 게놈에 의해 시퀀싱되었으며,여기에는 폐쇄 형 게놈이 포함됩니다. 지금까지 서열화 된 모든 미세 골 디아 균주의 게놈 특징은 표 1 에 요약되어 있습니다. 프로카를 사용한 주석은 게놈 당 1570~1906 개의 코딩 서열을 검출했으며,균주 당 평균 1760 개의 서열을 검출했다.
표 1 파인 골디아 균주의 서열화 된 게놈의 특징.
게놈 비교
에 따라 두 개의 서로 다른 클래드로 파인 골 디아 균주의 부문 우리는 이전에 파인 골 디아 분리체의 게놈 사이에서 큰 이질성을 발견했습니다 15. 모든 17 게놈의 핵심 게놈 정렬이 이루어졌고 핵심 게놈의 단일 뉴클레오티드 다형성이 호출되었습니다. 총,126,647 코어 게놈 스피 확인 하 고 계통 게놈 재건에 사용 되었다,공개 17 게놈 두 개의 서로 다른 클래드로 그룹화 될 수 있습니다(그림. 1). 코어 게놈의 평균 뉴클레오티드 식별(애니)을 결정하는 추가 분석은 두 개의 클래드가 90.7%의 애니,90.2%에서 91.2%로 변형 특이 적 애니 변이를 나타내는 것으로 나타났다. 이러한 낮은 애니는 두 개의 클래드가 개별 종 16 을 나타냄을 나타낸다. 마그나 클래드,96.6%(변이 95.1–97.4%)의 클래드 내 애니. 94.8%(93.4-96.이 균주는 다른 균주(균주 53516)와 주로 환자로부터 분리 된 균주를 포함합니다. 이후,이 계통의 균주는 잠정적으로”파인 골디아 네리 시아”(라틴어…에 대한 엔 ,이 종의 대부분의 균주가 분리 된 스웨덴 지역).
그림 1
계통 발생 트리 코어-게놈 모든 17 시퀀싱 된 게놈 의 파인 골디아 균주. 이 프로그램은 자바 바이트코드 프로그램의 갯수를 카운트하고,스크립트의 메인 형식을 합계냅니다,그리고 확인되지 않은 실행 텍스트 파일을 찾습니다.. 핵심 게놈은 참조 게놈의 43%로 표시됩니다. 게놈은 두 개의 주요 클래드에 할당 할 수 있습니다;하나의 클래드는 에프 마그나 균주를 나타내고 다른 클래드는 새로운 종의 균주로 구성되며 여기에서 잠정적으로”에프 네리 시아”.
16 초의 비교 르나 모든 분리 물의 서열은 높은 수준의 전반적인 유사성을 나타 냈지만(>99%),”에프 네리 시아”분리 물로부터 에프 마그나 분리 물의 분리를 검출했다. 그러나 이것은 매우 중요합니다. 또한 에프 마그나에 비해”에프 네리 시아”분리 물 사이에서 더 높은 다양 화를 나타 냈습니다. 다음,우리는 추가 분석 16. 이 경우,모든 것이 잘 작동합니다. 이것은 대부분의 저장된 시퀀스가”에프. 담즙 스텐트 생물막으로부터 분리 된 균주 17. 다시 말하지만,”에프 네리 시아”분리 물 중에서 더 높은 다양 화가 검출됩니다. 다음,우리 인간 마이크로바이옴 데이터(통합된 미생물 게놈 및 마이크로바이옴 샘플)데이터베이스에 저장 된 에프 마그나와”에프 네리 시아”시퀀스의 존재에 대 한 분석. (데이터는 표시되지 않음)에 존재하는 875 개의 조립된 인간 마이크로바이옴 데이터세트에서 식별될 수 있다. 이러한 분석을 종합하면 인간과 관련된 미세 골디아 분리체 중에서”에프 네리 시아”가 에프 마그나보다 더 널리 퍼진 것 같습니다.1362>피네골디아 속의 개체군 구조에 대해 더 자세히 밝히기 위해,12 개의 게놈(새로 서열화된 모든 10 개의 균주 및 2 개의 균주)의 비교 분석을 수행하였으며,여기에는 피네골디아 속의 4 개의 균주 및”피네골디아”의 8 개의 균주가 포함되었다. 12 게놈의 시각적 비교는 두 클레이드 사이의 뉴클레오티드 정체성 차이를 강조했습니다(그림 2). 2). 흥미롭게도,29328 에서 발견 된 플라스미드는 다른 서열 균주가 유사한 서열을 포함하지 않기 때문에이 균주에 대해 특이 적으로 보인다(그림 1). 2 에이).
그림 2
파인 골 디아 속에 속하는 12 가지 균주의 게놈 비교. 가장 안쪽 두 개의 고리는 지+씨-내용(검은 색)과 지씨-스큐(보라색/녹색)를 나타냅니다. 마그나);세 개의 가장 안쪽 게놈의 높은 뉴클레오티드 정체성(진한 붉은 색으로 시각화)을 참고(에프. (8 개의 외부 고리의 옅은 붉은 색);내부에서 외부로:균주 07,08,09,09,09,09,09,09,09,09,09,09,09,07,08,08,08,09,09,09,09,09,09,09,09,09,09,09,07,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08,08, 왼쪽 위 영역에 있는 플라스미드 29328 특정 영역은 플라스미드 29328 특정 영역을 나타냅니다. (2)기준 균주는 09 티 494(“에프. 7 개의”에프 네리 시아”게놈(내부 고리)은 4 개의 에프 마그나 게놈(외부 고리)에 비해 더 높은 뉴클레오티드 식별(어두운 붉은 색으로 표시);내부에서 외부로: 2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 12 월 1 일,2018 년 네리 시아”-특정 게놈 영역,하지만 5 개의 큰 스트레인 특정 클러스터를 참고.
게놈의 크기와 개수에 유의 한 차이는 두 클래드 사이에 검출되지 않았다. 또한,영창 분석(그림 1)에서 판단한 바와 같이 클레 드 특이 적(즉,에프 마그나 또는”에프 네리 시아”특이 적)게놈 영역이 검출되지 않았다. 2)그리고 또한 단백노르토(표 에스 1)를 사용하는 모든 핵분열의 양방향 폭발 접근법에 의해 확인되었다. 이 접근법은 모든 12 개의 게놈에 의해 공유되는 1202 개의 오르 로그,즉 각 균주의 평균 68%에서 핵심 프로테옴의 일부입니다. 단 아주 몇몇 클레이드 특정 유전자는 발견된다. 대신,단지 몇 개의 균주에 의해 공유되는 많은 영역이 발견될 수 있다. 2 비). 또한,변형률-특이 적 영역은 총 1016 개의 변형률-특이 적 변이체를 인코딩하는 것으로,균주 사이에 큰 범위가 존재한다. 균주 특이 적 기능 중에는 다양한 수송 기능(철,유출 펌프,올리고 펩타이드 수송 체),표면 단백질,제한 변형 시스템,접합 전달 기능,폴리 케타이드 합성,박테리오신,파지 관련 기능,항생제 내성 결정 인자 및 크리스퍼/카스 시스템이 있습니다.1362>흥미롭게도,크리스퍼/캐스 시스템의 차이는 에프 마그나와”에프 네리시아”의 균주들 사이에서 검출되었다. 구 에프 중 일곱. 마그나 균주는 균주 당 1~4 개의 크리스퍼 어레이를 포함했으며,평균 12 개의 스페이서(균주 당 3~38 개의 스페이서 범위)를 포함합니다. 반복”GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC”특정 변종의 F.magna. 반면,”에프 네리 시아”균주는 평균 53 스페이서(균주 당 14~87 범위)에 포함 된 균주 당 하나의 크리스퍼 어레이 만 수행했습니다. 각 변형은 변형률 별 이질성을 낮추고 각 변형에 고유 한 진화 역사를 가지고 있음을 나타내는 크리스퍼 배열의 개별 스페이서 세트를 포함합니다.
이러한 변형특성 함수가 수평적으로 획득되었는지를 평가하기 위해 아일랜드뷰어 프로그램을 사용하여 이동/획득의 징후를 조사했다. 이 프로그램은 예측 141(스트레인 09 티 408)408(스트레인 12 티 273)유전자는 수평 인수 될 가능성이 있는 게놈 섬에 있는,전문화에 관련 된 위에서 언급 한 스트레인 특정 유전자의 많은 포함 하 여,피트 니스,생존 및 종 간 경쟁(테이블 에스 3).
숙주 상호 작용 인자 및(추정)독성 인자
여러 숙주 상호 작용 인자가 설명되었다. 정렬 타제 의존성 필리는 접착 및 식민지화에 중요한 것으로 확인되었습니다 13. 모든 서열화 된 게놈 중에서 필루스 서브 유닛 에프엠 1,추정 서브 유닛 에프엠엠 2 및 3 개의 분류 효소에 대한 유전자를 암호화하는 게놈 궤적이 보존됩니다(그림 1). 3 에이). 이 궤적의 상향 및 하류 변형 특정 변형이 있습니다. 일부 균주는 헤 몰리 신 3 패밀리 단백질을 코딩하는 유전자를 포함하여 소르 타제 궤적의 상류 및 하류의 추가 유전자를 보유합니다. 흥미롭게도,에 관한 스트레인 별 차이가 있습니다. 거의 각 균주는 개별 변이체를 전달하며 단백질 수준에 대한 계통 발생 학적 분석은 핵심 게놈의 계통 발생 학적 분석과 상관 관계가 없었다(그림 1). 3 비). 대조적으로,에프엠피 2 는 모든 균주 중에서 매우 보존되어 있습니다. 콜라겐-및 피브리노겐 결합 도메인의 상류로 암호화 된 또 다른 단백질; 콜라겐 결합 도메인은 또한 콜라겐 결합 도메인에 존재합니다. 이 단백질은 또한 필루스 궤적에 속하며 해당 유전자의 이름을 지정합니다.
그림 3
소르 타제 의존성 필리 인코딩 게놈 유전자좌 파인 골디아 균주의 게놈에서. (에이)에 유전자좌의 게놈 조직 에프. 세 가지 분류 효소 유전자는 밝은 녹색으로 표시됩니다(“2″로 표시).; 주요 필루스 서브 유닛의 유전자는 적색(“1″이라고 표시됨)으로 표시되고,적색(“3″이라고 표시됨)은 주황색(“3″이라고 표시됨)으로 표시됩니다. 일부 균주에서는 추가 유전자(“8″,헤 몰리 신 3 가계 단백질 및”10”,가설 단백질)가 존재합니다. 콜라겐 결합 단백질(“4″로 표시)은 상류로 인코딩됩니다. (2)계통 발생 나무 1 파인 골디아 균주. 대부분의 균주는 개별 변종을 가지고 있습니다. 진화 역사는 메가 7 에서 수행 된 최대 우도 방법을 사용하여 추론되었습니다.
단백질 엘 아마 에프 마그나 8 의 가장 잘 연구 된 요인 일 것입니다. 면역 글로불린,면역 글로불린,면역 글로불린,면역 글로불린,면역 글로불린,면역 글로불린,면역 글로불린. 지금까지 테스트된 미세골디아 분리체의 약 10%에서 검출되었지만,크기가 다양하며,항원 결합 단편의 수가 4,8,18 이다. 변형 312 에서 연구 된 단백질 리터 폭발 검색(궤적 아이디:엠 86697,719 금주 모임)몇 가지 매우 유사한(단백질 정체성 50%이상)다양한 크기의 단백질(603 에 1417 금주 모임)몇 가지 균주에서,즉. 100,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000,000 4). 이 종에서 세포 초 항원이 더 널리 퍼져 있음을 나타냅니다. 또한 펩토스트렙토코커스 알부민 결합 단백질(아래 참조)로부터 알려진 2 개의 조지아 모듈을 보유하고 있다. 단백질 유사성이 낮은 많은 추가 단백질 엘,하지만 어떤 명백한 포함 하지 않고 비 1 도메인 게놈 서열 환 골 디아 균주에 인코딩됩니다.; 또,균주-특이 적 변이가 존재하고,몇몇 균주,즉 09 티토 408,09 티토 494 및 티토 151023 은 그러한 상동체를 갖지 않는다.
그림 4
유전자좌는 단백질 및 동족체를 미세골디아 균주에서 인코딩하는 유전자좌입니다. (1)유전체 서열화 된 미세 골디아 균주에서 수행되었으며,유사성이 높은(>50%단백질 정체성)동족체(적색,”1″로 표시됨)가 도시되었다(또한 표 참조). “에프 네리 시아”균주 만 에프는 아닙니다. 마그나 균주는 다양한 길이를 가진 그러한 동족체를 함유한다. (B)폭발 검색으로 공부 FAF 단백질의 변형 ALB8 찾을 전체 길이 homologs(에 빨간색 표시된”1”)에서 세 F. 마그나(빨간색)및 네”F.nericia”(밝은 파란색)종자. 이 유전자는 두 가지 균주 중 하나입니다.
파인골디아 마그나는 인간 알부민 9 에 결합하는 것으로 밝혀졌다. 펩토스트렙토코커스 알부민 결합 단백질은 균주 알부민 8 에서 연구되었다;그 활성은 가속 성장률과 관련이 있다 9. 균주 알 8 의 팹 단백질(궤적 이드: 19.이 모듈은 알부민 결합 도메인이 포함되어 있으며,조지아 모듈의 구조가 확인되었습니다. 폭발 검색은 가까운 팹 호모 로그가 다른 서열화 된 파인 골디아 게놈에서 발견 될 수 없다는 것을 밝혀 냈습니다. 대신,최대 50%의 단백질 정체성을 공유하는 몇 가지 상동체가 있습니다;에프 마그나 균주의 상동체의 유사성은”에프 네리 시아”균주의 상동체와 비교하여 평균적으로 더 높습니다. 가 모듈 함유 단백질에 관해서는 총 16 개의 단백질이 12 개의 미세 골디아 게놈에 암호화되어 알부민 결합 단백질의 일부 중복성을 나타냅니다. 대부분의 균주는 균주 09 를 제외하고 조지아 모듈로 두 개의 단백질을 암호화합니다.
세포벽 부착 서브 틸리 신 유사 프로테아제 수파는 콜라겐,피브리노겐 및 항균 펩타이드와 같은 여러 인간 단백질을 분해하는 것으로 나타났다. 폭발 검색은 대부분의 미세 골디아 균주가 변형 알 8 에서 연구 된 수파와 매우 유사한(단백질 정체성>75%)수파 호몰로그를 가지고 있음을 확인했습니다. 이 균주는 다음과 같습니다. 두 번째 수파 호몰로그는 대부분의 균주에 존재하며 평균 단백질 정체성은 26%입니다.
또 다른 중요한 호스트-상호 작용 인자는 다음과 같다. 마그나 균주의 약 90%가 이 인자를 생성하는데,이 인자는 박테리아의 응집을 담당하고 기저막에 결합함으로써 비엠-4011 에 결합한다. 우리는 7(3 에프 마그나와 4″에프 네리 시아”균주)12 균주 중 알 8 균주(그림. 4 비). 균주 특이 적 차이가 존재하며 프레임 시프트 돌연변이는”에프 네리 시아”균주 티 151023 과 12 티 272. 균주에서는 동족체를 찾을 수 없다.우리는 다른 추정 호스트 상호 작용 요인에 대 한 게놈을 검색 하 고 크리스티-앳 킨 스-뭉크-피터슨(캠프)요인에 대 한 코딩 유전자를 발견. 캠프 인자는 공동 헤 몰리 신으로 작용할 수 있으며 소위 캠프 반응,포도상 구균 아우 레 우스 스 핑고 미엘리나 제에 의한 양 적혈구의 상승 작용 용해 다(베타 독소)및 캠프 인자 22.
게놈 채굴은 모든 게놈 서열화 된 파인 골디아 균주에서 캠프 인자를 코딩하는 두 개의 유전자를 확인했다. 흥미롭게도,균주 당 하나의 호모 로그(캠프 2)는 추가적인 박테리아 유사 도메인을 보유합니다. 캠프 1 과 캠프 2 에 추가,네”에프. 네리 시아”균주는 다른 동족체와 실질적으로 다른 두 가지 추가 캠프 인자를 포함합니다(캠프 3 및 캠프 4;그림 에스 4). 따라서 7 개의”에프 네리 시아”균주 중 4 개는 4 개의 캠프 인자 유전자를 가지고 있습니다.
다음으로,우리는 파인 골디아 균주가 긍정적 인 캠프 반응을 일으키는지 알고 싶었습니다. 캠프 한천 플레이트 분석 결과 일부 균주는 강하게 캠프 반응 양성 동안 다른 균주에 대 한 단지 작은 공동 용혈 배양(그림 2)의 2 및 5 일에 적용 된 재배 조건 하에서 검출 될 수 있다 보여줍니다. 5,그림 5). 세 가지 테스트 에프. 마그나 균주는 강하게 캠프 반응 양성이었다. 두 균주는 강력 하 게 캠프 반응 양성,4 균주는 약간 공동 용혈성,그리고 한 균주는 명확 하 게 캠프 반응 음성. 유전체 분석 결과,균주 54800 에서 캠프 1-인코딩 유전자는 유전자의 5′-끝에 트랜스포사제-인코딩 궤적을 삽입하여 유전자를 교란시키는 것으로 나타났다(그림 6).
그림 5
파인 골 디아 균주의 캠프 반응. 양 혈액이 5%인 한천 판을 사용했습니다. 황색 포도상 구균은 중간 줄무늬에 접종됩니다. 양성 캠프 테스트는 미세 골디아의 인터페이스에서 완전한 적혈구 용해에 의해 표시됩니다. 아우 레 우스 줄무늬. 3 개의 마그나(빨간색)균주와 7 개의”에프 네리 시아”(파란색)균주 중 2 개는 혐기성 배양 2 일 후에 강한 양성 캠프 반응을 보였다.
우리의 결과는 그 캠프 요인을 나타냅니다. 기능적 일 수 있으며 캠프 요인 활동에 스트레인 별 차이가 있습니다.2 개의 생화확적인 시험 체계는”에프 네리키아”와 에프 마그나를 명확하게 구별할 수 없다. 설명 된;파인 골디아 사이에서만 약간의 생화학 적 변이. 이전에보고 된 균주 2,7. 우리는 혐기성 균의 식별에 사용 되는 두 개의 상업적으로 사용할 수 있는 생 화 확 적인 테스트 시스템을 테스트. 이 키트에는 21 개의 테스트가 포함되어 있으며 그 중 16 개의 탄수화물 활용 테스트가 포함되어 있습니다. 이전에보고 된 바와 같이 7,탄수화물 중 어느 것도 미세 골디 아에 의해 대사 될 수 없습니다. 변형(데이터는 표시되지 않음). 다른 반응(인돌,우레아제,카탈라아제,에스 쿨린 가수 분해 및 젤라틴 액화)은 또한 에프 마그나와”에프 네리 시아”균주 사이의 차이를 나타내지 않았다(데이터는 표시되지 않음). 이 키트에는 효소 활동에 대한 18 가지 테스트가 포함되어 있습니다. 당분 분해 효소가 부족한 반면,모든 미세골 디아. 균주는 단백질 분해 효소를 생산합니다. 포스파타제 활성을 검출하는 반응은 모든 에프 마그나 균주가 분명히 음성이고 대부분의”에프 네리 시아”균주가 양성인 균주 사이에서 다른 유일한 반응이었습니다. 그러나,이 시험 결과의 해석은 색깔 변화가 중요성의 경계에 있었기 때문에 한정됩니다. 결론적으로,기존의 두 가지 상용 생화학 검사 시스템은”에프 마그나”와”에프 네리 시아”를 명확하게 구별 할 수 없습니다. 미세 골 디아의 생화학 적 특성. 추가 생화확적인 시험으로 탐구될 필요가 있으십시오.
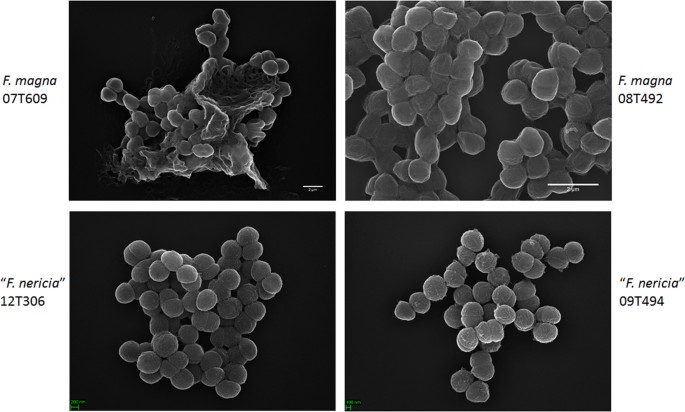
현미경 검사는 에프 마그나와”에프 네리 시아”의 형태와 표면 구조를 시각화하기 위해 에프 마그나와”에프 네리 시아”사이의 표면 차이를 보여준다. 그 결과,전자 현미경은 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 전자 현미경을 사용하여 마그나는 세포 외 고분자 매트릭스(그림 1)를 생산 겉으로는,더 부착 또는 집계했다. 6). 이것은 전자 현미경뿐만 아니라 공 초점 레이저 스캐닝 현미경 23 을 주사하여 생물막에서 엑소 다당류 매트릭스를 가시화 한 이전 작업을 확인합니다. 에서 테스트”에프. 네리 시아”균주 우리는 매트릭스를 식별 할 수 없습니다;세포는 덜 집계 보인다.
그림 6
그 결과,전자 현미경은 전자 현미경으로 검사됩니다. 또한,상기 패널들은 상기 패널들의 상부 및 하부 이미지들을 나타내고,상기 패널들은 상기 패널들의 상부 및 하부 이미지들을 나타내고 있다. 세포 에프 마그나 균주 더 접착 성/응집성 및 중합체 매트릭스를 생성하는 것,특히 균주 07 티 609;대조적으로,”에프 네리 시아”의 세포는 덜 응집성이다.
이러한 관찰을 확인했습니다. 또한,우리는 세포 부속에 관한 차이를 관찰했다. 마그나 세포는 전체 세포 표면에서 돌출 된 더 긴 필라멘트 부속기를 생성했다;이러한 구조는 필리 또는 핌 브리아를 나타낼 수 있습니다(그림 1). 7). 대조적으로,”에프 네리 시아”균주의 세포는 긴 돌출 된 부속기없이 더 매끄러운 표면을 가졌다. 그러나 그것은 배제할 수 없다,,이러한 무생물 관측은 세균 재배 조건 및 샘플 처리 단계에 따라 달라 집니다.
그림 7
원자력의 미세한 비교 에프 마그나 과”에프 네리 시아”. 왼쪽 패널과 오른쪽 패널은 각각 12 개의 이미지를 나타냅니다. (C)고해상도 이미지와 지원은 고급 소프트 터치 방식으로 Z 규모의 바이 최대 0.3μm XY 바을 나타내는 500nm. (비 과 디)의 고 대비 이미지의 오버레이 이미지(ㅏ 과 씨),독립적 인 수평 및 수직 다항식 제거와 결합(ㅏ 과 씨),11 도 해결. 실험은 각 균주로부터 3 개의 샘플에 대해 수행되었다.