- Inleiding
- Experiment 1
- methoden
- deelnemers
- apparaat en taak
- experimentele fasen en Procedure
- gegevensanalyse
- resultaten
- totale bewegingstijd
- relatieve Timing
- discussie
- Experiment 2
- methoden
- deelnemers
- apparatuur, taak, experimentele fasen, Procedure en gegevensanalyse
- Resultaten
- Totale bewegingstijd
- relatieve Timing
- discussie
- algemene discussie
- Auteursbijdragen
- financiering
- verklaring inzake belangenconflicten
- voetnoten
Inleiding
u bent een fervent golfer en u wilt een nieuwe opname leren. Hoe gaat u te werk? Er is een goede kans dat je iemand observeert (live, op video, op Youtube, enz.) wie weet hoe dit schot uit te voeren, en je zal proberen te begrijpen wat te doen en hoe het te doen. Onderzoek geeft duidelijk aan dat deze leerstrategie succesvol is omdat observatie is aangetoond dat het leren van een breed scala aan motorische vaardigheden wordt bevorderd (zie McCullagh et al., 1989; Hodges et al., 2007; Vogt and Thomaschke, 2007; Ste-Marie et al., 2012; Lago-Rodríguez et al., 2014, voor beoordelingen over observationeel leren). Dit komt omdat observatie veel gemeen heeft met fysieke oefening, die de eerste determinant is van motorisch leren. In het bijzonder is aangetoond dat variabelen, zoals de hoeveelheid praktijk (Carroll en Bandura, 1990; Blandin, 1994) , the frequency of knowledge of results (, Badets and Blandin, 2004, 2005; Badets et al., 2006), en het trainingsschema (Blandin et al., 1994; Wright et al., 1997), beïnvloeden leren via observationele praktijk en fysieke praktijk op soortgelijke manieren. Deze gegevens leidden tot de stelling dat observatie en fysieke praktijk zeer vergelijkbare processen gebruiken. Deze stelling wordt ondersteund door de resultaten van neuroimaging studies die aantoonden dat een ensemble van neurale structuren (met inbegrip van de premotorische cortex, de inferieure pariëtale lobule, de superieure temporale sulcus, de aanvullende motorische gebied, de cingulate gyrus, en de cerebellum), ook wel het “action observation network” (Aon) (Kilner et al., 2009; Oosterhof et al., 2010), wordt geactiveerd zowel wanneer individuen een bepaalde motorische taak uit te voeren en wanneer ze anderen die dezelfde motorische taak (Grafton et al., 1997; Buccino et al., 2001; Gallese et al., 2002; Cisek en Kalaska, 2004; Frey en Gerry, 2006; Cross et al., 2009; Dushanova and Donoghue, 2010; Rizzolatti and Fogassi, 2014; Rizzolatti et al., 2014).
observatie bevordert motorisch leren, maar wie moet je observeren om dat nieuwe golf schot te leren? Een expert die het schot waarschijnlijk beheerst zal u helpen een referentie te ontwikkelen van wat te doen en hoe het te doen, maar moet u iemand zoals u observeren die dat schot leert en die u vermoedelijk een betere kans geeft om fouten of veranderingen in strategie op te sporen en te leren? Onderzoek heeft aangetoond dat het observeren van zowel een bekwaam model (Martens et al., 1976; McCullagh et al., 1989; Lee et al., 1994; Al-Abood et al., 2001; Heyes and Foster, 2002; Hodges et al., 2003; Bird and Heyes, 2005) en een nieuw model leidt tot significant leren (Lee and White, 1990; McCullagh and Caird, 1990; Pollock and Lee, 1992; McCullagh and Meyer, 1997; Black and Wright, 2000; Buchanan et al., 2008; Buchanan and Dean, 2010; Hayes et al., 2010). Recente resultaten van ons laboratorium toonden echter aan dat het observationeel leren van een nieuwe motoriek verbeterd is na observatie van zowel beginnende als expertmodellen in plaats van een beginnend of een expertmodel alleen (Rohbanfard and Proteau, 2011; Andrieux and Proteau, 2013, 2014). Wij geloven dat dit “variabele” waarnemingsformaat niet alleen leidt tot de ontwikkeling van een goede bewegingsrepresentatie (deskundige observatie), maar ook tot de ontwikkeling van efficiënte processen voor foutdetectie en-correctie (beginnende observatie).
in deze studie is de kwestie van belang eenvoudig maar belangrijk. Bij het gebruik van een variabel observatieschema, zal het leren beter zijn wanneer de waarnemers vooraf worden geïnformeerd over de “kwaliteit” van de prestaties die ze op het punt staan te zien of zal het beter zijn wanneer de waarnemers worden overgelaten om de prestaties te evalueren alvorens feedback te ontvangen. Het informeren van de waarnemers van wat ze op het punt staan te zien kan hen in staat stellen om te kiezen of ze zullen observeren om te imiteren of liever observeren om fouten of zwakke punten in de prestaties van het model op te sporen, die de ontwikkeling van deze processen kunnen vergemakkelijken. Als alternatief, het hebben van de deelnemers evalueren van de prestaties kwaliteit die zij waargenomen kan meer uitgebreide cognitieve processen activeren dan wanneer deze informatie wordt ingevoerd (bijvoorbeeld, fout detectie en herkenning, of evaluatie van alternatieve strategie), wat resulteert in een beter leren van de taak.
de taak die we kozen vereiste dat de deelnemers het relatieve tijdspatroon veranderden dat natuurlijk voortkwam uit de taakbeperkingen (Collier and Wright, 1995; Blandin et al., 1999) tot een nieuw, opgelegd patroon van relatieve timing. Dit is vergelijkbaar met het veranderen van het tempo bij het uitvoeren van een serve in tennis of een drive in golf (Rohbanfard en Proteau, 2011). De deelnemers observeerden twee modellen die een grote verscheidenheid aan voorstellingen demonstreerden. In één groep werden de waarnemers vóór elke proef op de hoogte gebracht van het kwaliteitsniveau (prestaties van deskundigen, gevorderden, tussenpersonen, beginners of beginners) van wat zij zouden zien, terwijl een tweede groep waarnemers pas na afloop van elke proef dezelfde informatie kreeg.
Experiment 1
methoden
deelnemers
negentig rechtshandige studenten (45 mannen en 45 vrouwen; gemiddelde leeftijd = 20,5 jaar; SD = 0,9 jaar) uit het departement kinésiologie van de Université de Montréal namen aan dit experiment deel. De deelnemers waren naïef voor het doel van de studie en hadden geen eerdere ervaring met de taak, en alle deelnemers werden zelf verklaard als zijnde rechtshandig. Geen van de deelnemers meldde neurologische stoornissen en hadden allemaal een normaal of gecorrigeerd-naar-normaal gezichtsvermogen. De deelnemers hebben vóór de deelname individuele toestemmingsformulieren ingevuld en ondertekend. De Health Sciences Research Ethics Committee van de Université de Montréal heeft dit experiment goedgekeurd.
apparaat en taak
het apparaat was vergelijkbaar met dat van Rohbanfard and Proteau (2011). Zoals afgebeeld in Figuur 1, bestond het uit een houten voet (45 × 54 cm), drie houten barrières (11 × 8 cm) en een startknop ingebed in een doel (11 × 8 cm). De afstand tussen de startknop en de eerste slagboom was 15 cm. De afstanden van de overige drie segmenten van de taak waren respectievelijk 32, 18 en 29 cm. De barrières werden loodrecht op de houten basis geplaatst aan het begin van elke proef, wat een gesloten microschakelaar circuit opleverde. Alle microschakelaars werden aangesloten op een computer via de I / O-poort van een A-D converter (National Instruments, Austin, Texas, USA), en een milliseconde timer werd gebruikt om zowel de totale bewegingstijd (TMT) en de tijd die nodig is om elk segment van de taak te voltooien (tussenliggende tijden, ITs).
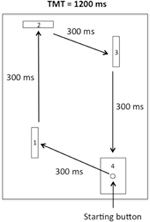
figuur 1. Schets van het apparaat. Deelnemers moesten de startknop verlaten en met de klok mee de eerste, tweede en derde slagboom raken voordat ze uiteindelijk het doel bereikten.
voor de praktijkproeven (zie hieronder) zaten de deelnemers dicht bij de uitgangspositie voor het apparaat. Vervolgens, vanaf de startknop, werden de deelnemers gevraagd om achtereenvolgens de eerste, tweede en derde barrières neer te halen (waardoor de microschakelaars worden losgelaten) en uiteindelijk het doel te raken in een beweging met de klok mee zoals geïllustreerd in Figuur 1. Elk segment van de taak moest worden voltooid in een IT van 300 ms, voor een TMT van 1200 ms. Het bewegingspatroon, zijn, en TMT werden geïllustreerd op een poster direct voor het apparaat tijdens alle experimentele fasen.
experimentele fasen en Procedure
de deelnemers werden willekeurig toegewezen aan een van de drie groepen, elk bestaande uit 30 deelnemers (15 vrouwen per groep): controle (C), feedforward KR en observatie (FW), en observatie en feedback KR (FB). Alle groepen voerden vier experimentele fasen uit, verdeeld over 2 opeenvolgende dagen.
Alle deelnemers kregen mondelinge instructies met betrekking tot TMT en ITs voor de eerste experimentele fase. De eerste experimentele fase was een pre-test, waarin alle deelnemers 20 physical practice trials uitvoerden zonder kennis van de resultaten (KR) op de TMT en de ITs.
de tweede fase was een acquisitiefase en bestond uit 60 observatieproeven voor de deelnemers in de twee observatiegroepen (FW en FB). Deze deelnemers keken individueel naar een videopresentatie van twee modellen die de experimentele taak fysiek uitvoerden. Voor elk observatieonderzoek werd KR met betrekking tot de prestaties van het model (zowel TMT als ITs) in ms gepresenteerd (zie Figuur 1), hetzij vóór de demonstratie voor de FW-groep, hetzij na de demonstratie voor de FB-groep. Het model werd om de vijf proeven gewijzigd (d.w.z., model 1: proeven 1-5 en model 2: proeven 6-10, enzovoort), voor een totaal van 30 proeven uitgevoerd door het ene model en 30 proeven uitgevoerd door het andere model. Voor zowel de FW-als FB-groepen werden de twee modellen, die deelnamen aan eerder werk vanuit ons laboratorium, gekozen omdat we voor beide modellen zes videoclips hadden die uitvoeringen in elk van de vijf subcategorieën illustreerden. Zo konden de deelnemers in de FW-en FB-groepen één bepaald model niet associëren met een betere of een slechtere prestatie. De prestaties van een expert kwamen overeen met een wortel gemiddelde vierkantfout (RMSE; zie de sectie gegevensanalyse voor rekendetails) variërend tussen 0 en 15 ms; de prestaties voor gevorderden, intermediairs, beginners en beginners kwamen overeen met RMSE ‘ s van respectievelijk 30-45 ms, 60-75 ms, 90-105 ms en 120+ ms. De deelnemers aan de FW-en FB-groepen werden op de hoogte gebracht van de prestaties van het model in de lidstaten; zij werden ook op de hoogte gebracht van het prestatieniveau waarnaar het verwijst. De resulterende 30 proeven van elk model (vijf prestatieniveaus × zes herhalingen) werden gerandomiseerd zodat de vijf prestatieniveaus eenmaal in elke reeks van vijf proeven werden gepresenteerd. Om fysieke imitatie van de sequentie te voorkomen, die de observationele processen zou kunnen verstoren, vroegen we de deelnemers aan de FW-en FB-groepen om hun handen op hun dijen te houden tijdens de acquisitiefase en om de bewegingen niet te reproduceren tijdens het bekijken van het model(de modellen). Het was de belangrijkste taak van de experimentator om ervoor te zorgen dat de deelnemers deze instructies opvolgden. Het openlijke gedrag van de deelnemers suggereert dat ze dat deden. Tenslotte hebben de deelnemers aan de controlegroep tijdens deze fase niets fysiek geoefend of geobserveerd. In plaats daarvan lezen ze een aangeboden krant of tijdschrift voor dezelfde duur als de observatie voor de andere groepen (ongeveer 10 min).
de derde en vierde experimentele fase waren 10-min en 24-uur retentiefasen. In elke fase, Alle deelnemers fysiek uitgevoerd 20 proeven zonder KR. De deelnemers werd gevraagd om elk segment van de taak in 300 ms te voltooien, voor een TMT van 1200 ms.
gegevensanalyse
de gegevens van de pre-test en de twee retentiefasen werden gegroepeerd in blokken van vijf onderzoeken. Voor elk volgend blok van vijf proeven (d.w.z., proeven 1-5, 6-10, enz.), berekenden we de absolute waarde van de constante fout van elke deelnemer (|CE|, de constante fout geeft aan of een deelnemer de totale bewegingstijd overschrijdt of overschrijdt) en variabele fout van de totale bewegingstijd (VE of binnen-deelnemer variabiliteit) om de nauwkeurigheid en consistentie van TMT, respectievelijk te bepalen. Voor tussenliggende tijden hebben we een RMSE berekend, die aangeeft hoeveel elke deelnemer in een enkele score afweek van het voorgeschreven relatieve tijdspatroon. Voor elke studie,
waarbij ITi de tussenliggende tijd voor segment “i” vertegenwoordigt en target de doelverplaatsingstijd voor elk segment van de taak (d.w.z. 300 ms).
omdat de gegevens normaal niet werden verspreid (RMSE en tijdgegevens zijn positief scheef), onderging elke afhankelijke variabele een logaritmische transformatie (ln). De getransformeerde gegevens voor elke afhankelijke variabele werden onafhankelijk ingediend bij een ANOVA die drie groepen (C, FW en FB) × drie fasen (pre-test, 10-min retentie, 24-h retentie) × vier blokken van de studies (1-5, 6-10, 11-15, en 16-20) met herhaalde metingen op de laatste twee factoren contrasteerde. Alle belangrijke hoofdeffecten en eenvoudige hoofdeffecten met meer dan twee middelen werden door de aanpassing van Bonferroni uitgesplitst. Voor alle vergelijkingen werd een effect significant geacht als p < 0,05. Partieel ETA-vierkant (np2) is de effectgrootte die gerapporteerd wordt voor alle significante effecten (Cohen, 1988).
resultaten
totale bewegingstijd
de ANOVA berekend op |CE| (Figuur 2, bovenpaneel) onthulde significante belangrijkste effecten voor de variabele Groep, F(2, 87) = 5,04, p = 0,08, np2 = 0,10, en fase, F(2, 174) = 5,16, p = 0,007, np2 = 0,06, evenals een significante fase × groepsinteractie, F(4, 174) = 4,93, p = 0,001, np2 = 0,10. De uitsplitsing van deze interactie liet geen significante groepsverschillen in de pre-test zien (f < 1). In de 10-min retentietest, F (2, 87) = 10,12, p < 0,001, np2 = 0,19, bleek uit de post-hoc vergelijkingen dat de controlegroep een significant groter | CE| had dan zowel de FW-als de FB-groepen (p < 0,05 in beide gevallen), die niet significant van elkaar verschilden (p = 0,19). In de 24-uur-retentietest, f (2, 87) = 4,34, p = 0,016, np2 = 0,09, had de FW-groep een significant kleiner |CE| dan de controlegroep (p = 0,012) 1.
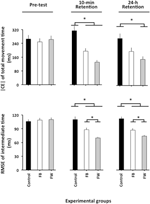
Figuur 2. Absolute constante fout van TMT en gemiddelde kwadraatfout van relatieve timing als functie van de experimentele fasen en experimentele groepen (Experiment 1). * p < 0,05. Foutbalken geven standaardfout van het gemiddelde aan.
de op VE berekende ANOVA(niet getoond) onthulde significante hoofdeffecten voor de variabele fase, F(2, 174) = 13,12, p < 0,001, np2 = 0,13, en block, F (3, 261) = 48,79, p < 0,001, np2 = 0,36. Post-hoc vergelijkingen van het faseeffect toonden een grotere VE van de totale tijd aan in de pre-test dan in zowel de 10-min als de 24-uur retentietests (p < 0.002 in beide gevallen), die niet significant van elkaar verschilden (p = 0,68). Het blok belangrijkste effect was het gevolg van een significant grotere VE van de totale tijd voor de eerste dan voor de drie resterende blokken van de studies (p < 0,001 in alle gevallen), die niet significant van elkaar verschilden (p>0,05 in alle gevallen).
relatieve Timing
de ANOVA berekend op de RMSE van relatieve timing onthulde significante belangrijkste effecten voor de variabele Groep, F(2, 87) = 21,49, p < 0,001, np2 = 0,33, fase, F(2, 174) = 39,98, p < 0,001, np2 = 0.31 en blok, F (3, 261) = 14,77, p < 0,001, np2 = 0,14, evenals een significante fase × groepsinteractie, F(4, 174) = 12,81, p < 0,001, np2 = 0,23. Het blok belangrijkste effect was het gevolg van een significant grotere RMSE van relatieve timing voor de eerste dan voor de drie resterende blokken van de studies (p < 0,001 in alle gevallen), die niet significant van elkaar verschilden (p>0,3 in alle gevallen). Wat interessanter is, is dat de uitsplitsing van de fase × groepsinteractie (Figuur 2, onderste paneel) geen significante groepsverschillen in de pre-test aan het licht bracht (F < 1). In de 10-min, F(2, 87) = 14,85, p < 0,001, np2 = 0,34, en de 24-h-retentietesten, F(2, 87) = 23,23, p < 0,001, np2 = 0,35, hoewel de FB-groep significant beter presteerde dan de controlegroep (p = 0,001 in beide gevallen), was de FB-groep op zijn beurt significant beter dan de FW-groep (P = 0,001 en P = 0,02, respectievelijk)2.
discussie
dit experiment was bedoeld om onze kennis van de observatieomstandigheden uit te breiden die het leren van een nieuw relatief tijdspatroon optimaliseren. In deze leersituatie werden twee observatiegroepen, die een verscheidenheid aan demonstraties waarnamen, voor of na elke proef tijdens de acquisitiefase KR verstrekt. In het bijzonder wilden we beoordelen of het leren zou worden verbeterd als de leerlingen de “kwaliteit” of kenmerken van een demonstratie kennen voordat ze de demonstratie observeren. De resultaten zijn eenvoudig.
Ten eerste presteerden zowel de FW-als de FB-groepen beter dan de controlegroep bij de retentietests. Dit gold voor het leren van zowel de TMT als de relatieve timing. Dit verwachte resultaat bevestigt eerdere bevindingen die erop wezen dat observatie iemand in staat stelt om een nieuwe motorische vaardigheid te leren (zie McCullagh et al., 1989; Hodges et al., 2007; Vogt and Thomaschke, 2007; Ste-Marie et al., 2012; Lago-Rodríguez et al., 2014, for reviews on observational learning) en, in het bijzonder, een nieuw relatieve tijdspatroon (Rohbanfard and Proteau, 2011; Andrieux and Proteau, 2013, 2014).
de belangrijkste bevinding van dit onderzoek is dat de FB-groep beter presteerde dan de FW-groep in de retentietests. Hoewel de twee groepen dezelfde demonstraties waarnamen, bleek uit de resultaten dat het leren geoptimaliseerd is wanneer men vooraf kennis krijgt van de kwaliteit of kenmerken van de getuige demonstratie. Deze bevinding sluit goed aan bij eerdere rapporten van ons laboratorium (Rohbanfard and Proteau, 2011; Andrieux and Proteau, 2013) waaruit blijkt dat een gemengd observatieregime, waarin de waarnemers weten wie het expertmodel is en wie het beginnersmodel is, het leren van een nieuw relatief tijdspatroon beter begunstigt dan deskundige of beginnende observatie alleen.
van tevoren weten dat een minder dan perfecte demonstratie zal worden getoond kan van cruciaal belang zijn, gezien het feit dat is gemeld dat beginnende deelnemers, zoals in deze studie, niet goed zijn in het beoordelen van de kwaliteit van een demonstratie. Bijvoorbeeld, Aglioti et al. (2008) had beginnende en deskundige basketbalspelers observeren videoclips met vrije worp schoten, en de videoclips werden gestopt op verschillende tijdstippen voor of onmiddellijk na de bal release. Deskundige basketbalspelers en coaches / gespecialiseerde journalisten waren beter en sneller in het voorspellen van het lot van het schot (succesvol of niet) dan waren beginners (voor soortgelijke resultaten zie ook Wright et al., 2010; Abreu et al., 2012; Tomeo et al., 2013; Balser et al., 2014; Candidi et al., 2014; Renden et al., 2014).
het voordeel van de FW ten opzichte van het FB-protocol is belangrijk en, voor zover bekend, is een soortgelijke bevinding tot nu toe niet gerapporteerd. Daarom leek een replicatie van deze bevinding belangrijk. Bovendien vroegen we ons af of het voor het FW-protocol genoteerde voordeel pas na een beperkte hoeveelheid observatie optrad. Tot slot waren we benieuwd of het afwisselen van het FW en het FB protocol tot additieve effecten zou leiden. We voerden Experiment 2 uit om deze vragen aan te pakken.
Experiment 2
methoden
deelnemers
de 60 deelnemers die vrijwillig aan dit experiment deelnamen, waren afkomstig uit dezelfde populatie als die van Experiment 1 (36 mannen en 24 Vrouwen; gemiddelde leeftijd = 22,7 jaar; SD = 4,9 jaar). De deelnemers waren naïef over het doel van deze studie en hadden geen eerdere ervaring met de taak. Zij hebben vóór de deelname individuele toestemmingsformulieren ingevuld en ondertekend. De Health Sciences Research Ethics Committee van de Université de Montréal heeft dit experiment goedgekeurd.
apparatuur, taak, experimentele fasen, Procedure en gegevensanalyse
we gebruikten dezelfde taak, apparatuur en procedures als in Experiment 1. Het grote verschil tussen het huidige experiment en Experiment 1 is dat de deelnemers twee acquisitiesessies uitvoerden, wat leidde tot een totaal van vijf experimentele fasen: pre-test, acquisitie 1, onmiddellijke retentietest, acquisitie 2, en 24-h retentietest.
de deelnemers werden willekeurig toegewezen aan een van de drie groepen, elk bestaande uit 20 deelnemers (8 vrouwen per groep): feedforward KR en observatie tijdens zowel acquisitie 1 en 2 (FW1-2); feedforward observatie en KR tijdens acquisitie 1 maar observatie en feedback KR tijdens acquisitie 2 (FW/FB); en observatie en KR feedback tijdens zowel acquisitie 1 en 2 (FB1-2). We gebruikten dezelfde video en modellen als in Experiment 1; echter, de volgorde van de videopresentatie was anders in acquisitie 2 van die in acquisitie 1. Alle deelnemers werden ook geïnformeerd dat ze dezelfde taak zouden uitvoeren na elke acquisitiefase, maar zonder KR met betrekking tot hun eigen prestaties.
we gebruikten dezelfde afhankelijke variabelen en datatransformatie als in Experiment 1. Voor elke afhankelijke variabele hebben we een ANOVA in twee richtingen uitgevoerd waarin de drie groepen (FW1-2, FW/FB en FB1-2) × drie experimentele fasen (pre-test, onmiddellijke retentie en 24-uur retentie) werden vergeleken. Alle belangrijke hoofdeffecten en eenvoudige hoofdeffecten met meer dan twee middelen werden door de aanpassing van Bonferroni uitgesplitst. Voor alle vergelijkingen werd een effect significant geacht als p < 0,05. Partieel ETA-vierkant (np2) is de effectgrootte die gerapporteerd wordt voor alle significante effecten (Cohen, 1988).
Resultaten
Totale bewegingstijd
De ANOVA berekend voor het |CE| verkeer (Figuur 3) bleek een significante hoofdeffecten voor de variabele groep, F(2, 57) = 8.13, p = 0.001, np2 = 0.22, en fase, F(2, 114) = 21.13, p < 0.001, np2 = 0.27, evenals een aanzienlijke groep × fase interactie, F(4, 114) = 2.57, p = 0.042, np2 = 0.08. De uitsplitsing van deze interactie liet geen significante groepsverschillen in de pre-test zien (f < 1). In de onmiddellijke retentietest, F (2, 57) = 10,27, p < 0,002, np2 = 0.De FB1-2-groep had een significant groter| CE / dan zowel de FW1-2-als de FW/FB-groepen (p < 0,001 in beide gevallen), die niet significant van elkaar verschilden (p>0,20). In de 24-h-retentietest, F (2, 57) = 3,19, p = 0,049, np2 = 0,10, had de FW1-2-groep een iets kleiner| CE / dan de FB1-2-groep (p = 0,079)3.
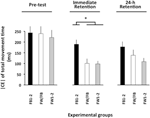
Figuur 3. Absolute constante fout van TMT als functie van de experimentele fasen en experimentele groepen (Experiment 2). * p < 0,05. Foutbalken geven standaardfout van het gemiddelde aan.
de ANOVA berekend op VE (niet getoond) onthulde significante hoofdeffecten voor de variabele Groep, F(2, 57) = 7,82, p = 0,001, np2 = 0,21, en fase, F(2, 114) = 21,10, p < 0,001, np2 = 0,27, evenals een significante groep × fase interactie, F(4, 114) = 4,38, p = 0,002, np2 = 0,13. De uitsplitsing van deze interactie liet geen significante groepsverschillen zien in de pre-test (F < 1) en in de 24-uur-retentietest, F(2, 57) = 1,26, p>0,20. In de onmiddellijke retentietest, F (2, 57) = 10,26, p < 0,002, np2 = 0,27, de FB1-2-groep (62.7 ms) had een significant grotere VE dan zowel de FW1-2 (51,1 ms) als de FW/FB (53,4 ms) groepen (p < 0,001 in beide gevallen), die niet significant van elkaar verschilden (p>0,20)4.
relatieve Timing
de ANOVA die werd berekend voor de RMSE van relatieve timing onthulde significante hoofdeffecten voor de variabele Groep, F(2, 57) = 4,86, p = 0,01, np2 = 0,15, en phase, F(2, 114) = 78,21, p < 0,001, np2 = 0,58. Er was een significant grotere RMSE van relatieve timing in de pre-test dan in zowel de onmiddellijke-retentietest als de 24-uur-retentietest (p < 0.001 in beide gevallen; zie Figuur 4, rechterpaneel), die niet significant van elkaar verschilden (p>0,20). Ten slotte presteerden de FW1-2 en de FW/FB groepen beter dan de FB1-2 groep (p = 0,01 en p = 0,07; zie Figuur 4, linkerpaneel) maar verschilden niet significant van elkaar (p>0,20).
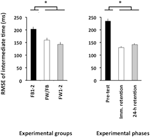
Figuur 4. Root gemiddelde kwadraatfout van relatieve timing (Experiment 2) als functie van de experimentele groepen (linkerpaneel) en experimentele fasen (rechterpaneel). * p < 0,05. Foutbalken geven standaardfout van het gemiddelde aan.
discussie
zoals verwacht, ondersteunt de afname in fouten die werd waargenomen bij het overgaan van de pre-test naar de retentietests eerdere bevindingen die erop wijzen dat observatie helpt bij het leren van een nieuw relatieve tijdspatroon (Blandin et al., 1999; Rohbanfard and Proteau, 2011; Andrieux and Proteau, 2013, 2014). Belangrijker, repliceerden de resultaten van Experiment 2 die van Experiment 1, in die zin dat de groep FW1-2 de groep FB1-2 overtrof. Daarom kan veilig worden geconcludeerd dat het leren om het relatieve tijdspatroon te veranderen dat van nature uit de beperkingen van de taak naar een nieuwe, opgelegde relatieve tijd door observatie komt, de voorkeur krijgt wanneer men wordt geïnformeerd over de prestaties van het model voorafgaand aan in plaats van na observatie. Tot slot toonden de resultaten ook aan dat wat geleerd is in een FB-protocol niet optelt tot wat geleerd kan worden in een FW-protocol.
algemene discussie
het hoofddoel van deze studie was om te bepalen wanneer in een observatieprotocol KR betreffende de modelprestaties moet worden verstrekt, d.w.z. vóór of na elke demonstratie. Uit de resultaten van de twee experimenten van deze studie blijkt duidelijk dat de kennis van de prestaties van het model vóór elke demonstratie het leren van een nieuw relatieve tijdspatroon beter beviel dan wanneer de waarnemer na elke demonstratie op de hoogte werd gesteld van de prestaties van het model. Bovendien suggereren de resultaten van Experiment 2 dat het voordeel van de FW ten opzichte van het FB-protocol significant bleef, zelfs wanneer het aantal observatieproeven werd verdubbeld. Met betrekking tot dit laatste punt stellen wij niet dat een FW-protocol in alle gevallen en met alle niveaus van deskundigheid van de waarnemers moet worden begunstigd. Integendeel, we benadrukken dat het effect betrouwbaar is wanneer beginnende waarnemers worden beschouwd.
onze resultaten kunnen erop wijzen dat een feedforward observation protocol de waarnemer voorbereidt om specifiek deel te nemen aan ofwel imitatieprocessen wanneer een deskundige of geavanceerde prestatie wordt getoond of in foutdetectieprocessen wanneer een beginner of beginnende prestatie wordt gepresenteerd. Dit idee past goed bij vorig werk van Decety et al. (1997), die stelde dat de patronen van hersenactivering tijdens actieobservatie afhangen van zowel de aard van de vereiste uitvoerende verwerking als de extrinsieke eigenschappen van de gepresenteerde actie. Specifiek, deze auteurs toonden aan dat verschillende gebieden van de hersenen actiever worden wanneer men observeert te herkennen, wat het geval zou kunnen zijn bij het observeren van een beginner model of een slechte of intermediaire prestaties, en wanneer men observeert te imiteren, wat waarschijnlijk het geval is bij het observeren van een expert model.
een alternatieve verklaring voor onze bevindingen zou kunnen zijn dat een FW-protocol resulteert in een “deactivering” van de AON wanneer de deelnemers expliciet werden geïnformeerd dat er een slechte demonstratie zou volgen. Bijvoorbeeld, in een object-lifting taak, is aangetoond dat de modulatie van motor evoked potential (MEP) door transcraniële magnetische stimulatie (TMS) tijdens de observatie van de lifting actie wordt geschaald naar de kracht die nodig is om de greep en lifting actie uit te voeren (Alaerts et al., 2010a). Er werd ook aangetoond dat wanneer visuele aanwijzingen suggereerden dat het object zwaarder was dan in werkelijkheid was, de MEP-modulatie voornamelijk afhankelijk was van het waargenomen kinematische profiel in plaats van van het schijnbare gewicht van het object (Alaerts et al., 2010b; Senot et al., 2011). Echter, in een studie van Senot et al. (2011), valse expliciete informatie over het gewicht van het object werd verstrekt in een experimentele toestand. Dit resulteerde in een conflict tussen het verwachte kinematische profiel gezien het aangekondigde gewicht en het werkelijke kinematische profiel van de grijp-en hijswerking, wat leidde tot een “algemene remming van het corticospinale systeem.”Anders gesteld, ten minste een deel van de AON was uitgeschakeld. Het kan daarom zijn dat de deelnemers aan ons onderzoek de AON hebben uitgezet toen slechte prestaties van het model werden verwacht, waardoor de AON alleen actief bleef voor goede proeven.
deze stelling is echter moeilijk te verzoenen met recente rapporten van ons laboratorium waaruit blijkt dat het observeren van zowel een expert-als een beginnersmodel resulteerde in een beter leren van een nieuw relatieve tijdspatroon dan het observeren van een beginnersmodel of een expertmodel alleen. Als men de AON zou kunnen uitschakelen wanneer men zou weten dat er een slechte demonstratie zal worden getoond (d.w.z. een beginnersmodel), dan zou het leren van de gemengde observatiegroep hebben geëvenaard en niet overtroffen dat van de deskundige observatiegroep. In plaats daarvan, terug te gaan naar onze eerste propositie, stellen we voor dat een FW-protocol helpt beginnende performers detecteren en kwantificeren van fouten in de prestaties van het model, iets wat ze meestal slecht doen (Aglioti et al., 2008; Wright et al., 2010; Abreu et al., 2012; Tomeo et al., 2013; Balser et al., 2014; Candidi et al., 2014; Renden et al., 2014). Op zijn beurt kan de betere detectie en kwantificering van de prestaties van het model de ontwikkeling van inverse (Jordan, 1996) en forward modellen (Wolpert en Miall, 1996) van motor control bevorderen.
concluderend kan worden gesteld dat observatie een krachtig leermiddel is dat voor iedereen beschikbaar is en dat slechts minimale apparatuur vereist om te worden gebruikt. Het is nu goed aangetoond dat de voordelen van observatie voor het wijzigen van de relatieve timing (dat wil zeggen, tempo) van motorische vaardigheid worden versterkt wanneer men toegang heeft tot een verscheidenheid van prestaties, variërend van beginners tot deskundigen, hetzij door middel van variabele of gemengde observatie schema ‘ s. De resultaten van deze studie suggereren dat deze voordelen geoptimaliseerd worden als de waarnemer vooraf de kwaliteit kent van de prestaties die hij of zij tijdens de eerste observatiesessie gaat waarnemen. Dit kan erg belangrijk zijn in een klaslokaal context waarin een leraar/trainer een video observatie protocol zou gebruiken. Bijvoorbeeld, als de bedoeling van de waarnemer is om een specifiek aspect van een golf swing te leren, is het waarschijnlijk dat het resultaat van de swing (dat wil zeggen, de bal vlucht) zal niet worden weergegeven op de video. Daarom zou de waarnemer niet in staat zijn om de expertise van het model te” raden ” aan de hand van het resultaat van de swing en, zoals we in deze studie hebben aangetoond, beter te leren of hij of zij van tevoren was geïnformeerd over de kwaliteit van wat hij of zij op het punt staat te observeren.
Auteursbijdragen
alle genoemde auteurs hebben een aanzienlijke, directe en intellectuele bijdrage aan het werk geleverd en het voor publicatie goedgekeurd.
financiering
dit werk werd ondersteund door een Discovery Grant (LP) van de Natural Sciences and Engineering Research Council of Canada (grant no. 111280-2013).
verklaring inzake belangenconflicten
de auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.
voetnoten
1. ^Om vast te stellen dat de verschillen die in de twee retentietesten tussen de controlegroep en de FW en FB-groepen werden geconstateerd, het gevolg waren van een significante afname van de totale bewegingstijd, hebben we in een aanvullende analyse de groep × fase-interactie die in de hoofdtekst werd gerapporteerd, ontleed door voor elke groep een aparte ANOVA te berekenen. Uit de resultaten is gebleken dat Voor de controle-en de FB-groepen het |CE| van de totale verplaatsingstijd niet significant verschilde tussen de fasen, respectievelijk. Integendeel, voor de FW-groepen was er een significant hoofdeffect van de fasen, F (2, 86) = 11.60, p < 0,01, np2 = 0,1, waaruit een significante afname van de| CE / van de totale bewegingstijd bleek, gaande van de pre-test naar de twee retentietests (p < 0,01), die niet significant van elkaar verschilden (p>0,10).
2. ^Zoals we deden voor de / CE / van de totale bewegingstijd, ontleden we in een aanvullende analyse de groep × fase interactie gerapporteerd in de hoofdtekst voor de RMSE van relatieve timing door het berekenen van een aparte ANOVA voor elke groep. De resultaten toonden aan dat Voor de controlegroep de RMSE van relatieve timing niet significant verschilde tussen de fasen, F(2, 86) = 0,32, p = 0,72, np2=0,01. Integendeel, zowel voor de FB-als voor de FW-groep was er een significant hoofdeffect van de fasen dat een significante afname van de RMSE van relatieve timing aantoonde, gaande van de pre-test naar de twee retentietests (p < 0,01), die niet significant van elkaar verschilden (p > 0,10).
3. ^Zoals we deden in Experiment 1, in een aanvullende analyse ontleden we de groep × fase interactie gerapporteerd in de hoofdtekst door het berekenen van een aparte ANOVA voor elke groep. Uit de resultaten bleek dat Voor de FB1-2-groep het| CE / van de totale bewegingstijd niet significant verschilde tussen de fasen, F(2, 56) < 1, p = 0,45, np2 = 0,03. Integendeel, voor zowel de FW1-2-als de FW-FB-groepen was er een significant hoofdeffect van de fasen, dat Voor beide groepen een significante afname van de totale verplaatsingstijd van de pre-test naar de twee retentietests (p < 0) aan het licht bracht.01), die niet significant van elkaar verschilden (p>0,10).
4. ^Voor de VE van de totale bewegingstijd bleek uit de uitsplitsing van de groep × fase-interactie een significant hoofdeffect van fasen voor alle drie groepen . Voor de FW1-2-en de FB1-2-groepen bleek uit pos-hoc vergelijkingen een significant grotere VE in de pre-test dan in beide retentietests (p < 0,01), die niet significant van elkaar verschilden (p>0,30). Voor de FW-FB-groep was de VE van de totale bewegingstijd significant groter in de pre-test dan in de 24-uur-retentietest (p < 0,01).
Abreu, A. M., Macaluso, E., and Azevedo, R. T. (2012). Actievoorspelling buiten het action observation network: een functionele magnetic resonance imaging studie bij deskundige basketbalspelers. Euro. J. Neurosci. 35, 1646–1654. doi: 10.1111 / j. 1460-9568. 2012. 08104.X
PubMed Abstract / CrossRef Full Text / Google Scholar
Aglioti, S. M., Cesari, P., Romani, M., and Urgesi, C. (2008). Actie anticipatie en motorische resonantie bij elite basketbalspelers. Nat. Neurowetenschappen. 11, 1109–1116. doi: 10.1038 / nn.2182
PubMed Abstract / CrossRef Full Text / Google Scholar
Al-Abood, S. A., Davids, K. F., and Bennett, S. J. (2001). Specificiteit van taakbeperkingen en effecten van visuele demonstraties en verbale instructies bij het sturen van het zoeken van leerlingen tijdens het verwerven van vaardigheden. J. Mot. Behav. 33, 295–305. doi: 10.1080/00222890109601915
PubMed Abstract / CrossRef Full Text / Google Scholar
Alaerts, K., Senot, P., Swinnen, S. P., Craighero, L., Wenderoth, N., en Fadiga, L. (2010a). De krachtvereisten van waargenomen objectlifting worden gecodeerd door het motorsysteem van de waarnemer: een TMS-studie. Euro. J. Neurosci. 31, 1144–1153. doi: 10.1111 / j. 1460-9568. 2010. 07124.X
PubMed Abstract / CrossRef Full Text / Google Scholar
Alaerts, K., Swinnen, S. P., en Wenderoth, N. (2010b). Observeren hoe anderen lichte of zware objecten optillen: welke visuele signalen bemiddelen de codering van spierkracht in de primaire motorische cortex? Neuropsychologia 48, 2082-2090. doi: 10.1016 / j.neuropsychologia.2010.03.029
PubMed Abstract / CrossRef Full Text / Google Scholar
Andrieux, M., and Proteau, L. (2013). Observatie leren van een motorische taak: wie en wanneer? Exp. Brain Res. 229, 125-137. doi: 10.1007 / s00221-013-3598-X
PubMed Abstract / CrossRef Full Text / Google Scholar
Andrieux, M., and Proteau, L. (2014). Gemengde observatie bevordert motorisch leren door een betere schatting van de prestaties van het model. Exp. Brain Res. 232, 3121-3132. doi: 10.1007 / s00221-014-4000-3
PubMed Abstract / CrossRef Full Text / Google Scholar
Badets, A., and Blandin, Y. (2004). De rol van kennis van resultaten frequentie in het leren door middel van observatie. J. Mot. Behav. 36, 62–70. doi: 10.3200 / JMBR.36.1.62-70
PubMed Abstract / CrossRef Full Text / Google Scholar
Badets, A., and Blandin, Y. (2005). Observationeel leren: effecten van bandbreedte kennis van resultaten. J. Mot. Behav. 37, 211–216. doi: 10.3200 / JMBR.37.3.211-216
PubMed Abstract | CrossRef Full Text | Google Scholar
Badets, A., Blandin, Y., Wright, D. L., and Shea, C. H. (2006). Error detection processes during observational learning. Res. Q. Exerc. Sport 77, 177–184. doi: 10.1080/02701367.2006.10599352
PubMed Abstract | CrossRef Full Text | Google Scholar
Balser, N., Lorey, B., Pilgramm, S., Naumann, T., Kindermann, S., Stark, R., et al. (2014). De invloed van expertise op de hersenactivering van het actieobservatienetwerk tijdens het anticiperen op tennis en volleybal dient. Voorkant. Brom. Neurowetenschappen. 8:568. doi: 10.3389 / fnhum.2014.00568
PubMed Abstract / CrossRef Full Text / Google Scholar
Bird, G., and Heyes, C. (2005). Effector-afhankelijk leren door observatie van een vingerbeweging sequentie. J. Exp. Psychol. Brom. Percept. Uitvoeren. 31, 262–275. doi: 10.1037/0096-1523.31.2.262
PubMed Abstract / CrossRef Full Text / Google Scholar
Black, C. B., and Wright, D. L. (2000). Kan observationele praktijk foutherkenning en beweging productie vergemakkelijken? Re. Q. Oefening. Sport 71, 331-339. doi: 10.1080/02701367.2000.10608916
PubMed Abstract / CrossRef Full Text / Google Scholar
Blandin, Y. (1994). Cognitieve processen betrokken bij het leren van Spatio-temporele synchronisatie taken onder verschillende omstandigheden van de praktijk en observatie van een Model Subject, ongepubliceerde proefschrift. Universiteit van Montreal.
Blandin, Y., Lhuisset, L., en Proteau, L. (1999). Cognitieve processen die ten grondslag liggen aan observationeel leren van motorische vaardigheden. Q. J. Exp. Psychol. 52, 957–979. doi: 10.1080/713755856
CrossRef Full Text / Google Scholar
Blandin, Y., Proteau, L., en Alain, C. (1994). Over de cognitieve processen die ten grondslag liggen aan contextuele interferentie en observationeel leren. J. Mot. Behav. 26, 18–26. doi: 10.1080/00222895.1994.9941657
PubMed Abstract / CrossRef Full Text / Google Scholar
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L., Gallese, V., et al. (2001). Actieobservatie activeert premotorische en pariëtale gebieden op somatotopische wijze: een fMRI-studie. Euro. J. Neurosci. 13, 400–404. doi: 10.1046 / j. 1460-9568. 2001. 01385.X
PubMed Abstract / CrossRef Full Text / Google Scholar
Buchanan, J. J., and Dean, N. J. (2010). Specificiteit in de praktijk is gunstig voor leren in beginnende modellen en variabiliteit in demonstratie is gunstig voor observationele praktijk. Psychol. 74, 313-326. doi: 10.1007 / s00426-009-0254-y
PubMed Abstract / CrossRef Full Text / Google Scholar
Buchanan, J. J., Ryu, Y. U., Zihlman, K., and Wright, D. L. (2008). Observationele praktijk van relatieve maar niet absolute beweging kenmerken in een enkel-ledemaat multi-gezamenlijke coördinatie taak. Exp. Brain Res. 191, 157-169. doi: 10.1007 / s00221-008-1512-8
PubMed Abstract / CrossRef Full Text / Google Scholar
Candidi, M., Sacheli, L. M., Mega, I., and Aglioti, S. M. (2014). Somatotopic mapping of piano fingering errors in sensorimotor experts: TMS studies bij pianisten en visueel getrainde muzikaal naïven. Cereb. Cortex 24, 435-443. doi: 10.1093 / cercor / bhs325
PubMed Abstract / CrossRef Full Text / Google Scholar
Carroll, W. R., and Bandura, A. (1990). Representationele begeleiding van actieproductie in observationeel leren: een causale analyse. J. Mot. Behav. 22, 85–97. doi: 10.1080/00222895.1990.10735503
PubMed Abstract / CrossRef Full Text / Google Scholar
Cisek, P., and Kalaska, J. F. (2004). Neurale correlaten van mentale repetitie in de dorsale premotorische cortex. Nature 431, 993-996. doi: 10.1038 / nature03005
PubMed Abstract / CrossRef Full Text / Google Scholar
Cohen, J. (1988). Statistische Vermogensanalyse voor de gedragswetenschappen. Hillsdale, NJ: Lawrence Erlbaum.
Google Scholar
Collier, G. L., and Wright, C. E. (1995). Temporele herschaling van monster en complexe rantsoenen in ritmische tikken. J. Exp. Psychol. Brom. Percept. Uitvoeren. 21, 602–627. doi: 10.1037/0096-1523.21.3.602
PubMed Abstract / CrossRef Full Text / Google Scholar
Cross, E. S., Kraemer, D. J. M., Hamilton, A. F. D., Kelley, W. M., and Grafton, S. T. (2009). Gevoeligheid van het action observation network voor fysiek en observationeel leren. Cereb. Cortex 19, 315-326. doi: 10.1093 / cercor / bhn083
PubMed Abstract / CrossRef Full Text / Google Scholar
Decety, J., Grèzes, J., Costes, N., Perani, D., Jeannerod, M., Procyk, E., et al. (1997). Hersenactiviteit tijdens observatie van acties-invloed van actieinhoud en strategie van het onderwerp. Brain 120, 1763-1777. doi: 10.1093 / brain / 120.10.1763
PubMed Abstract / CrossRef Full Text / Google Scholar
Dushanova, J., and Donoghue, J. (2010). Neuronen in primaire motorische cortex ingeschakeld tijdens actie observatie. Euro. J. Neurosci. 31, 386–398. doi: 10.1111 / j. 1460-9568. 2009. 07067.X
PubMed Abstract / CrossRef Full Text / Google Scholar
Frey, S. H., and Gerry, V. E. (2006). Modulatie van neurale activiteit tijdens observationeel leren van acties en hun sequentiële orders. J. Neurosci. 26, 13194–13201. doi: 10.1523 / JNEUROSCI.3914-06.2006
PubMed Abstract / CrossRef Full Text / Google Scholar
Gallese V., Fadiga L., Fogassi L., Rizzolatti G. (2002). “Action representation and the inferior parietal lobule,” in Common Mechanisms in Perception and Action: Attention and Performance, Vol. XIX, eds W. Prinz, and B. Hommel (Oxford: Oxford University Press), 247-266.
Grafton, S. T., Fadiga, L., Arbib, M. A., and Rizzolatti, G. (1997). Promotor cortex activering tijdens observatie en naamgeving van bekende tools. Neuroimage 6, 231-236. doi: 10.1006 / nimg.1997.0293
PubMed Abstract / CrossRef Full Text / Google Scholar
Hayes, S. J., Elliott, D., and Bennett, S. J. (2010). Algemene motorische representaties worden ontwikkeld tijdens de actie-observatie. Exp. Brain Res. 204, 199-206. doi: 10.1007 / s00221-010-2303-6
PubMed Abstract / CrossRef Full Text / Google Scholar
Heyes, C. M., and Foster, C. L. (2002). Motorisch leren door observatie: bewijs van een seriële reactietijdstaak. Q. J. Exp. Psychol. 55, 593–607. doi: 10.1080/02724980143000389
PubMed Abstract / CrossRef Full Text / Google Scholar
Hodges, N. J., Chua, R., and Franks, I. M. (2003). De rol van video in het faciliteren van perceptie en actie van een nieuwe coördinatiebeweging. J. Mot. Behav. 35, 247–260. doi: 10.1080/00222890309602138
PubMed Abstract / CrossRef Full Text / Google Scholar
Hodges, N. J., Williams, A. M., Hayes, S. J., and Breslin, G. (2007). Wat wordt gemodelleerd tijdens observationeel leren? J. Sports Sci. 25, 531–545. doi: 10.1080/02640410600946860
PubMed Abstract / CrossRef Full Text / Google Scholar
Jordan, M. I. (1996). “Computational aspects of motor control and motor learning,” in Handbook of Perception and Action: Vol. 2, Motor Skills, eds H. Heuer en S. W. Keele (New York, NY: Academic Press), 71-120.
Kilner, J. M., Neal, A., Weiskopf, N., Friston, K. J., and Frith, C. D. (2009). Bewijs van spiegelneuronen in menselijke inferieure frontale gyrus. J. Neurosci. 29, 10153–10159. doi: 10.1523 / JNEUROSCI.2668-09.2009
PubMed Abstract / CrossRef Full Text / Google Scholar
Lago-Rodríguez, A., Cheeran, B., en Koch, G. (2014). De rol van spiegelneuronen in observationeel motorisch leren: een integratieve beoordeling. Euro. J. Human Mov. 32, 82–103.
Lee, T. D., Swinnen, S. P., and Serrien, D. J. (1994). Cognitieve inspanning en motorisch leren. Quest 46, 328-344. doi: 10.1080/00336297.1994.10484130
CrossRef Full Text / Google Scholar
Lee, T. D., And White, M. A. (1990). Invloed van een ongeschoolde modellen oefenschema op observationeel motorisch leren. Brom. Mov. Sci. 9, 349–367. doi: 10.1016/0167-9457(90)90008-2
CrossRef Full Text / Google Scholar
Martens, R., Burwitz, L., en Zuckerman, J. (1976). Modellering effecten op motor-prestaties. Res. Q. 47, 277-291.
PubMed Abstract / Google Scholar
McCullagh, P., and Caird, J. K. (1990). Correcte en leermodellen en het gebruik van modelkennis van resultaten in het verwerven en behouden van een motorische vaardigheid. J. Human Mov. Stud. 18, 107–116.
McCullagh, P., and Meyer, K. N. (1997). Leren versus correcte modellen: invloed van het modeltype op het leren van een vrijgewichtkraaklift. Res.Q. Exerc. Sport 68, 56-61. doi: 10.1080/02701367.1997.10608866
PubMed Abstract / CrossRef Full Text / Google Scholar
McCullagh, P., Weiss, M. R., And Ross, D. (1989). Modellering overwegingen in motorische vaardigheid acquisitie en prestaties: een geïntegreerde aanpak. Exerc. Sport Sci. Rev. 17, 475-513.
PubMed Abstract / Google Scholar
Oosterhof, N. N., Wiggett, A. J., Diedrichsen, J., Tipper, S. P., and Downing, P. E. (2010). Oppervlakte-gebaseerde informatie mapping onthult crossmodale visie-actie representaties in menselijke pariëtale en occipitotemporale cortex. J. Neurophysiol. 104, 1077–1089. doi: 10.1152 / jn.00326.2010
PubMed Abstract / CrossRef Full Text / Google Scholar
Pollock, B. J., and Lee, T. D. (1992). Effecten van het vaardigheidsniveau van het model op observationeel motorisch leren. Res.Q. Exerc. Sport 63, 25-29. doi: 10.1080/02701367.1992.10607553
PubMed Abstract / CrossRef Full Text / Google Scholar
Renden, P. G., Kerstens, S., Oudejans, R. R. D., and Cañal-Bruland, R. (2014). Fout of duik? Motorische bijdragen aan het beoordelen van dubbelzinnige foute situaties in het voetbal. Euro. J. Sport Sci. 14 (Suppl. 1), S221–S227. doi: 10.1080/17461391.2012.683813
PubMed Abstract / CrossRef Full Text / Google Scholar
Rizzolatti, G., Cattaneo, L., Fabbri-Destro, M., en Rozzi, S. (2014). Corticale mechanismen die ten grondslag liggen aan de organisatie van doelgerichte acties en spiegel neuron-gebaseerde actie begrip. Fysiol. Rev. 94, 655-706. doi: 10.1152 / physrev.00009.2013
PubMed Abstract / CrossRef Full Text / Google Scholar
Rizzolatti, G., and Fogassi, L. (2014). Het spiegelmechanisme: recente bevindingen en perspectieven. Philos. Transvetzuren. R. Soc. Lond. Ser. B Biol. Sci. 369:20130420. doi: 10.1098 / rstb.2013.0420
PubMed Abstract / CrossRef Full Text / Google Scholar
Rohbanfard, H., and Proteau, L. (2011). Leren door observatie: een combinatie van deskundige en beginnende modellen bevordert leren. Exp. Brain Res. 215, 183-197. doi: 10.1007 / s00221-011-2882-X
PubMed Abstract / CrossRef Full Text / Google Scholar
Senot, P., D ‘ ausilio, A., Franca, M., Caselli, L., Craighero, L., en Fadiga, L. (2011). Effect van gewichtsgerelateerde labels op corticospinale prikkelbaarheid tijdens observatie van grijpen: een TMS-studie. Exp. Brain Res. 211, 161-167. doi: 10.1007 / s00221-011-2635-X
PubMed Abstract / CrossRef Full Text / Google Scholar
Ste-Marie, D. M., Law, B., Rymal, A. M., Jenny, O., Hall, C., and McCullagh, P. (2012). Observatie interventies voor motorische vaardigheden leren en prestaties: een toegepast model voor het gebruik van observatie. Int. Rev.Sport Exerc. Psychol. 5, 145–176. doi: 10.1080 / 1750984X.2012.665076
CrossRef Full Text / Google Scholar
Tomeo, E., Cesari, P., Aglioti, S. M., and Urgesi, C. (2013). De kickers voor de gek houden, maar de Keepers niet.: gedrag en neurofysiologische correlaten van nep actie detectie in Voetbal. Cereb. Cortex 23, 2765-2778. doi: 10.1093 / cercor / bhs279
PubMed Abstract / CrossRef Full Text / Google Scholar
Vogt, S., and Thomaschke, R. (2007). Van visuo-motorische interacties tot imitatieleren: gedrags-en hersenbeeldvormingsstudies. J. Sports Sci. 25, 497–517. doi: 10.1080/02640410600946779
PubMed Abstract / CrossRef Full Text / Google Scholar
Wolpert, D. M., and Miall, R. C. (1996). Voorwaartse modellen voor fysiologische motorische controle. Neuraal Netwerk. 9, 1265–1279. doi: 10.1016/S0893-6080(96)00035-4
PubMed Abstract / CrossRef Full Text / Google Scholar
Wright, D. L., Li, Y., and Coady, W. (1997). Cognitieve processen gerelateerd aan contextuele interferentie en observationeel leren: een replicatie van Blandin, Proteau and Alain (1994). Res.Q. Exerc. Sport 68, 106-109. doi: 10.1080/02701367.1997.10608872
PubMed Abstract / CrossRef Full Text / Google Scholar
Wright, M. J., Bishop, D. T., Jackson, R. C., and Abernethy, B. (2010). Functional MRI reveals expert-novice differences during sport-related anticipation. Neuroreport 21, 94–98. doi: 10.1097/WNR.0b013e328333dff2
PubMed Abstract | CrossRef Full Text | Google Scholar