- Finegoldian kantojen genomit
- Finegoldian kantojen jakaminen kahteen erilliseen kladiin genomivertailun
- Finegoldia SP: n Kannaspesifinen heterogeenisuus
- isännän vuorovaikutustekijät ja (oletetut) virulenssitekijät
- Finegoldian genomit koodaavat Camp-tekijöitä, jotka ovat funktionaalisia
- kaksi biokemiallista testijärjestelmää ei voi yksiselitteisesti erottaa F. Magnaa ”F. nericiasta”
- mikroskopia paljastaa pinnan erot F. Magnan ja”F. nerician ”välillä
Finegoldian kantojen genomit
Finegoldian 17 genomia. analysoitiin ja verrattiin. Ryhmässämme 15 on hiljattain sekvensoitu kymmenen kantaa, jotka eristettiin ortopedisten nivelimplanttien aiheuttamista infektioista kärsiviltä potilailta Ruotsissa. Seitsemän genomia oli aiemmin sekvensoitu muilla, mukaan lukien tyypin kannan ATCC 2932814 suljettu genomi. Kaikkien finegoldian tähän mennessä sekvensoitujen kantojen genomipiirteet on esitetty taulukossa 1. Prokan avulla tehdyissä merkinnöissä havaittiin 1570-1906 koodaussekvenssejä (CDS) per genomi, keskimäärin 1760 CD per kanta.
Finegoldian kantojen jakaminen kahteen erilliseen kladiin genomivertailun
perusteella havaitsimme aiemmin Finegoldian isolaattien genomien keskuudessa suuren heterogeenisyyden 15. Kaikkien 17 genomin ydin-genomi-linjaus tehtiin ja perusgenomin yksittäisiä nukleotidipolymorfismeja (SNP) kutsuttiin. Yhteensä, 126,647 core-genomi SNPs tunnistettiin ja käytettiin fylogenomic rekonstruktio, paljastaen, että 17 genomit voidaan ryhmitellä kahteen eri kladeja (Fig. 1). Lisäanalyysi, joka määrittää ytimen genomin keskimääräisen nukleotiditunnuksen (ANI), osoitti, että kahden kladin ANI on 90,7%, ja kannan spesifinen ANI vaihtelu 90,2%: sta 91,2%: iin. Tällainen alhainen ANI osoittaa, että nämä kaksi kladia edustavat yksittäisiä lajeja16. Yksi kladi sisältää eniten aiemmin sekvensoituja ja tutkittuja kantoja, mukaan lukien ATCC29328 ja ALB8; sitä voidaan pitää klassisena F. magna kladena, jonka intraclade ANI on 96,6% (muunnelmat 95,1–97,4%). Toinen kladi, jonka intraklade ANI on 94, 8% (93, 4–96.6%) sisältää toista ATCC-kantaa (ATCC 53516) ja pääasiassa kantoja, jotka on eristetty ruotsalaisen Örebron yliopistollisen sairaalan potilaista15. Tämän kladin kantoja kutsutaan alustavasti nimellä ”Finegoldia nericia” (latinaksi Närke, Ruotsin alue, jossa suurin osa tämän lajin kannoista oli eristyksissä).

Fylogenominen puu, joka perustuu finegoldia-kantojen Kaikkien 17 sekvensoidun genomin ydin-genomeihin. Ohjelma Parstnp: n avulla linjattiin ytimen perimä ja kutsuttiin SNPs: ksi. Perusgenomia edustaa 43% referenssigenomista (ATCC29328); koko genomin fylogenian rekonstruoimiseen käytettiin yhteensä 126 647 luotettavaa perusgenomisidonnaista SNP: tä. Genomit voidaan jakaa kahteen pääkladiin; toinen kladi edustaa F. magna-kantoja ja toinen kladi koostuu jonkin uuden lajin kannoista, tässä alustavasti ”F. nericia”.
kaikkien isolaattien 16S rRNA-sekvenssien vertailu osoitti yleistä samankaltaisuutta (>99%), mutta havaitsi F. magna-isolaattien erottumisen ”F. nericia” – isolaateista, koska 5-9 ”F. nericia ” – erityiset kansalliset SNP: t (Kuva S1). Lisäksi ”F. nericia” – isolaattien keskuudessa havaittiin suurempi hajautus verrattuna F. Magnaan. Seuraavaksi analysoimme lisää 16s rRNA-sekvenssejä, jotka kuuluvat Finegoldia sp: lle. tallennettu Genbankiin (Kuva S2). Tämä paljasti, että useimmat tallennetut sekvenssit klusteroituvat ”F. nerician” kanssa, erityisesti ryhmän Finegoldia sp. sapen stentistä biofilms17 eristetyt kannat. Jälleen ”F. nericia” – isolaattien keskuudessa havaitaan suurempaa hajaantumista. Seuraavaksi analysoimme ihmisen mikrobiomitiedot, jotka on tallennettu img (Integrated Microbial Genomes and microbiome samples) – tietokantaan F. magna-ja ”F. nericia” – sekvenssien esiintymisestä. 3287 ja 7068 F. magna ATCC29328: n ja ”F. nericia” ATCC 53516: n koodaussekvenssit voitiin tunnistaa 875 ihmisen mikrobiomiaineistosta, jotka olivat IMG: ssä (tietoja ei näy); Finegoldian suurimmat esiintymät havaittiin retroaurikulaarisissa prässinäytteissä. Yhdessä nämä analyysit viittaavat siihen, että ihmiseen liittyvistä Finegoldia-isolaateista ”F. nericia” näyttää olevan yleisempi kuin F. magna.
Finegoldia SP: n Kannaspesifinen heterogeenisuus
Finegoldia-suvun populaatiorakenteen selvittämiseksi tehtiin vertailevia analyyseja 12 genomista (kaikki kymmenen uutta sekvensoitua kantaa ja kaksi ATCC-kantaa), mukaan lukien neljä F. magna clade-kantaa ja kahdeksan ”F. nericia” – kladin kantaa. 12 genomin visuaalinen vertailu korosti kahden kladin välistä nukleotidi-identiteettieroa (Kuva. 2). Mielenkiintoista on, että ATCC 29328: sta löydetty plasmidi näyttää olevan spesifinen tälle kannalle, koska mikään muu sekvensoitu kanta ei sisällä samanlaista sekvenssiä (Kuva. 2 A).

Genomivertailu 12: sta ”Finegoldia” – sukuun kuuluvasta kannasta. Kaksi sisintä rengasta edustavat G + C-pitoisuutta (musta) ja GC-vinoa (violetti/vihreä). (A) vertailukanta on ATCC29328 (F. magna); huomaa kolmen sisimmän genomin (jotka kaikki kuuluvat F: ään) suuri nukleotidi-identiteetti (jota havainnollistaa tummanpunainen väri). magna), ja alempi tunnistaa kahdeksasta ”F. nericia” – sukuun kuuluvasta kannasta (kahdeksan ulkorenkaan vaaleanpunainen väri); sisältä ulos: kannat 07t609, 08t492, 09t408, 09T494, 12t272, 12t273, 12t306, CCUG 54800, T151023, T160124 ja ATCC 53516. Suuri atcc29328-spesifinen alue vasemmassa yläkulmassa edustaa plasmidin pFMC: tä. B) vertailukanta on 09T494 (”F. nericia”). Seitsemällä” F. nericia ” – genomilla (Sisärenkaat) on korkeampi nukleotidi-tunniste (jota edustaa tummempi punainen väri) verrattuna neljään F. magna-genomiin (ulkorenkaat); sisältä ulospäin: kannat 12T272, ATCC 53516, 12T306, 12t273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 ja 09t408. ”F. nericia”-spesifisistä genomialueista ei ole merkkejä, mutta huomaa viisi suurta lajikohtaista klusteria.
näiden kahden kladin välillä ei havaittu merkittäviä eroja genomin koossa ja CD-levyjen määrässä. Myöskään kladi-spesifisiä (eli F. magna tai ”F. nericia” – spesifisiä) genomialueita ei havaittu, kuten priki-analyysissä arvioitiin (Kuva. 2) ja myös vahvistettu kaksisuuntainen blast lähestymistapa kaikkien CDS käyttäen ProteinOrtho (taulukko S1). Tässä lähestymistavassa havaittiin 1202 ortologiaa, jotka ovat yhteisiä kaikille 12 genomille, eli keskimäärin 68% kunkin kannan CD-arvoista on osa ydinproteomia. Kladispesifisiä geenejä löytyy vain hyvin vähän. Sen sijaan voidaan löytää monia alueita, joilla esiintyy vain muutamia kantoja, esimerkiksi kannalle 09T494 (Katso myös kuva. 2b). Lisäksi esiintyy kantakohtaisia alueita, jotka koodaavat yhteensä 1016 kantakohtaista CDS: ää, ja kantojen välinen vaihtelu on suuri, esimerkiksi 187 ja 22 kantakohtaista CDS: ää havaitaan kannoissa ATCC29328 ja T151023 (taulukko S1). Kantakohtaisia funktioita ovat muun muassa erilaiset kuljetusfunktiot (rauta, effluksipumput, oligopeptidien ABC-transportterit), pintaproteiinit, restriktiojärjestelmät, konjugatiiviset siirtofunktiot, polyketidisynteesi, bakteriosiinit, faageihin liittyvät funktiot, antibioottiresistenssin taustatekijät ja CRISPR/cas-järjestelmät.
mielenkiintoista on, että CRISPR/cas-järjestelmissä havaittiin eroja F. Magnan ja ”F. nerician” kantojen välillä (taulukko S2). Seitsemän yhdeksästä F: stä. magna-kannoissa oli yhdestä neljään CRISPR-ryhmää per kanta, ja niissä oli keskimäärin 12 välilevyä (3-38 välilevyä per kanta). Toistuva ”GTTTGAGAATGATGTATTAAC” oli spesifinen F. Magnan kannoille. Sen sijaan ”F. nericia” – kannoilla oli vain yksi CRISPR-ryhmä per kanta, joka sisälsi keskimäärin 53 välilevyä (vaihteluväli 14-87 per kanta). Jokainen kanta sisälsi CRISPR-ryhmässään (- ryhmissään) yksilöllisen välilevyjen joukon, joka alleviivaa kantakohtaista heterogeenisyyttä ja osoittaa, että kullakin kannalla on ainutlaatuinen evolutionaarinen historia.
arvioidaksemme, onko tällaisia kantakohtaisia toimintoja hankittu horisontaalisesti, tarkastelimme merkkejä niiden liikkuvuudesta / hankinnasta IslandViewer-ohjelman avulla (kuva S3). Tämä ohjelma ennusti 141 (kanta 09T408)-408 (kanta 12T273) geenejä, jotka sijaitsevat genomisaarilla ja jotka todennäköisesti hankitaan horisontaalisesti, mukaan lukien monet edellä mainituista lajikohtaisista geeneistä, jotka osallistuvat erikoistumiseen, kuntoiluun, selviytymiseen ja lajien väliseen kilpailuun (taulukko S3).
isännän vuorovaikutustekijät ja (oletetut) virulenssitekijät
useita isännän vuorovaikutustekijöitä on kuvattu F. magna 4,5: lle. Sortaasista riippuvaisia pilejä on tunnistettu, jotka ovat tärkeitä tarttumisen ja kolonisaation kannalta 13. Genominen lokus, joka koodaa geenit pilus alayksikkö Fmp1, putatiivinen alayksikkö Fmp2 ja kolme sortaasit on säilynyt kaikkien sekvensoitu genomit (Fig. 3 A). Tämän lokuksen ylä-ja alajuoksulla on kantakohtaisia variaatioita, ts. joillakin kannoilla on ylimääräisiä geenejä fmp1: n yläjuoksulla ja sortaasi-lokuksen alajuoksulla, mukaan lukien hemolysiini III-perheen proteiinia koodaava geeni. Mielenkiintoista on, että fmp1: n suhteen on kantakohtaisia eroja. Lähes jokaisella kannalla on yksilöllinen variantti, ja fmp1: n fylogeneettinen analyysi proteiinitasolla ei osoittanut korrelaatiota perusgenomin fylogenomisen analyysin kanssa (Fig. 3b). Sen sijaan Fmp2 säilyy hyvin kaikkien kantojen joukossa. Toinen proteiini, joka on koodattu ennen fmp1: tä, sisältää kollageenia ja fibrinogeeniä sitovia domeeneja; kollageenia sitova domeeni (COG4932) esiintyy myös fmp2: ssa. Ehdotamme, että tämä proteiini kuuluu myös pilus locus ja nimi vastaava geeni fmp3.

Sortaasiriippuvainen pili-koodaava genomiloktio Finegoldian kantojen genomeissa. (A) loci: n Genomijärjestö F. magna (punainen) ja ”F. nericia” (Vaaleansininen) – kannoissa. Kolme sortaasi-geeniä on kuvattu vaaleanvihreällä (merkitty ”2”); suuren pilus-alayksikön fmp1 geeni on merkitty punaisella (merkitty ”1”) ja fmp2 oranssilla (merkitty ”3”). Fmp1: n yläjuoksulla ja sortaasigeenien alajuoksulla on joissakin kannoissa muita geenejä (merkitty ”8”, hemolysiini III-perheen proteiini ja ”10”, hypoteettinen proteiini). Oletettu kollageenia sitova proteiini, Fmp3 (merkitty ”4”), koodataan ennen fmp1: tä. B) Finegoldia-kantojen Fmp1: n fylogeneettinen puu. Useimmilla kannoilla on yksilöllinen Fmp1-muunnos. Evoluutiohistoria on päätelty Mega7: ssä tehdyn Maksimitodennäköisyysmenetelmän avulla.
proteiini L on luultavasti parhaiten tutkittu tekijä F. magna 8. Se on superantigeeni, jonka kohteena ovat B-solut ja immunoglobuliinit. Sitä on havaittu noin 10%: ssa tähän mennessä testatuista Finegoldia-isolaateista, mutta sen koko ja antigeenia sitovien fragmenttien (Fab) määrä vaihtelee (B1-domeeni, pfam02246)4,8,18. Blast-haku tutkittavalla proteiinilla l kannasta 312 (lokus ID: M86697, 719 aa) paljastaa muutaman hyvin samankaltaisen proteiinin (proteiinin identiteetti yli 50%), joiden koko vaihtelee (603-1417 aa) muutamissa kannoissa, ts. kannat CCUG54800, T161024, ATCC53516, 12T273 ja 12t306 (Kuva. 4 A). Kaikki nämä ovat” F. nericia ” -kantoja, mikä viittaa siihen, että B-solun superantigeeni on yleisempi tällä lajilla. Poikkeuksellisen suurella (1417 aa) proteiinin l kaltaisella tekijällä kannassa 12t306 on kahden B1-domeenin lisäksi myös kaksi GA-modulia, jotka tunnetaan peptostreptokokin albumiinia sitovista proteiineista (KS.alla). Genomisekvensoituneisiin Finegoldia-kantoihin on koodattu monia muita proteiineja, joilla on alhainen samankaltaisuus proteiini L: n kanssa, mutta jotka eivät sisällä mitään näennäistä B1-domeenia (taulukko S4A); tässäkin lajikohtaisia variaatioita on, ja muutamilla kannoilla eli 09t408, 09T494 ja T151023 ei ole tällaisia homologeja.

genomiset loci koodaavat proteiini L: n ja FAF: n homologeja Finegoldian kannoissa. (A) Blastitutkimukset, joissa käytettiin kantaa 312 (lokuksen tunnus: m86697) olevaa tyypillistä proteiinia l, tehtiin genomisekvensoiduista Finegoldia-kannoista, ja homologeja (punaisella, merkintä ”1”), jotka ovat hyvin samankaltaisia (>50% proteiinin identiteetistä) on esitetty (KS.myös taulukko S4A). Ainoastaan” F. nericia ” – kantoja, mutta ei F. magna-kannat sisältävät tällaisia homologeja eripituisina. B) Blastitutkimuksissa, joissa käytettiin tutkittavaa kannan ALB8 FAF-proteiinia, löytyi täysmittaisia homologeja (punaisella, merkintä ”1”) kolmesta F. magna (punainen) – ja neljästä ”F. nericia” (Vaaleansininen) – kannasta. FAF-geeni on kehyksellinen kahdessa” F. nericia ” – kannassa T151023 ja 12t272.
Finegoldia Magnan havaittiin sitoutuvan ihmisen albumiini9. Alb8-kannassa on tutkittu peptostreptokokin aiheuttamaa albumiinia sitovaa proteiinia (PAB), jonka aktiivisuus on yhteydessä nopeutuneeseen kasvuvauhtiin9. Kannan ALB8 PAB-proteiini (locus ID: X77864) sisältää GA-moduulit, albumiinia sitovan domeenin; GA-moduulin rakenne on selvitetty19. Blast-haku paljasti, että läheistä PAB-homologia ei löydy muista sekvensoiduista Finegoldian genomeista (taulukko S4B). Sen sijaan on olemassa useita homologeja, joilla on jopa 50% proteiinidentiteetti; F. magna-kantojen homologien yhtäläisyydet ovat keskimäärin suuremmat kuin ”F. nericia” – kantojen homologien. GA-moduulia sisältävien proteiinien osalta 12 Finegoldia-genomiin koodataan yhteensä 16 proteiinia, mikä viittaa albumiinia sitovien proteiinien redundanssiin (taulukko S4C). Useimmat kannat koodaavat kahta proteiinia GA-moduuleilla, paitsi kannoista 09t408 (F. magna) ja T151023 (”F. nericia”).
soluseinään kiinnittyneen subtilisiinin kaltaisen proteaasin SufA on osoitettu hajottavan useita ihmisen proteiineja, kuten kollageenia IV,fibrinogeeniä ja antimikrobisia peptidejä kuten LL-37 ja MIG/CXCL910,20, 21. Blastitutkimuksissa todettiin, että useimmilla Finegoldian kannoilla on sufa-homolog, joka on hyvin samankaltainen (proteiinidentiteetti > 75%) kuin tutkittu SufA-kanta ALB8 (lokus ID: DQ679960). Poikkeuksia ovat kannat 07T609, 09T408 ja CCUG54800. Toinen SufA-homolog esiintyy useimmissa kannoissa; sen keskimääräinen proteiinidentiteetti on 26% (taulukko S4d).
toinen tärkeä isännän vuorovaikutustekijä on FAF (F. magna adheesiokerroin). Noin 90% F. magna-kannoista tuottaa tätä tekijää, joka on vastuussa bakteerien paakkuuntumisesta ja välittää solukalvoon sitoutumisen BM-4011: een. Havaitsimme, että seitsemän (kolme F. magna-kantaa ja neljä ”F. nericia” – kantaa) 12: sta kannasta koodaa FAF-homologeja, jotka muistuttavat tutkittua ALB8-kantaa (Kuva. 4b). Lajikohtaisia eroja on ja ”F. nericia” – kannoissa T151023 ja 12t272 nähdään kehyksensiirtomutaatioita. Homologiaei esiinny kannoissa 09T408, 09t494 ja 12t306 (taulukko S4E).
Finegoldian genomit koodaavat Camp-tekijöitä, jotka ovat funktionaalisia
etsimme genomista muita oletettuja isäntävoimatekijöitä ja löysimme Christie-Atkins-Munch-Petersen (CAMP) – tekijöitä koodaavia geenejä. CAMP-tekijät voivat toimia ko-hemolysiineinä ja selittää niin sanotun CAMP-reaktion, Staphylococcus aureus sphingomyelinase C: n (beetatoksiini) aiheuttaman lampaiden erytrosyyttien synergistisen lyysin ja CAMP-tekijän22.
Genominlouhinta tunnisti kaksi geeniä, jotka koodaavat CAMP-tekijöitä jokaisessa Genomisekvenssoidussa Finegoldian kannassa (CAMP1 ja CAMP2; Kuva S4); jokaisessa homologissa on CAMP-tekijäperhe (PFAM07373). Mielenkiintoista on, että yksi homolog (CAMP2) kantaa kohden sisältää ylimääräisen bakteerien Ig: n kaltaisen domeenin (Pfam02368). CAMP1: n ja CAMP2: n lisäksi neljä ”F. nericia ” – kannoissa on kaksi muuta CAMP-tekijää, jotka eroavat huomattavasti muista homologeista (CAMP3 ja CAMP4; Kuva S4). Niinpä neljällä seitsemästä” F. nericia ” – kannasta on neljä CAMP factor-geeniä.
seuraavaksi halusimme tietää, tuottavatko Finegoldia-kannat positiivisen CAMP-reaktion. CAMP agar-levymääritys osoittaa, että jotkin kannat ovat voimakkaasti CAMP-reaktiopositiivisia, kun taas muiden kantojen osalta voidaan havaita vain vähän koehemolyysiä sovelletuissa viljelyolosuhteissa kahden ja viiden päivän inkubaation jälkeen (kuva. 5, Kuva S5). Kaikki kolme testattua F. magna-kannat olivat vahvasti CAMP-reaktiopositiivisia. ”F. nericia” -kannoista havaitsimme jonkin verran heterogeenisuutta; kaksi kantaa oli voimakkaasti CAMP-reaktiopositiivisia, neljä kantaa vain hieman koehemolyyttisiä ja yksi kanta, CCUG54800, oli selvästi CAMP-reaktio-negatiivinen. Genomianalyysissä havaittiin, että kannassa CCUG54800 CAMP1-koodaava geeni sisälsi transposaasia koodaavan lokuksen geenin 5′-päähän, mikä häiritsee geeniä (Kuva S6).

Finegoldia-kantojen CAMP-reaktio. Käytettiin TSA agar-levyjä, joissa oli 5% lampaanverta. Staphylococcus aureus rokotetaan keskimmäisessä viirussa. Positiivinen CAMP-testi osoitetaan täydellisellä erytrosyyttilyysillä Finegoldia SP: n rajapinnassa. ja S. aureus juovuttaa. Kaikki kolme F. magna (punaisena) – kantaa ja kaksi seitsemästä ”F. nericia” (sinisenä) – kannasta osoittivat voimakkaan positiivisen CAMP-reaktion kahden päivän anaerobisen inkubaation jälkeen.
tuloksemme osoittavat, että FINEGOLDIA sp: n CAMP-tekijät. voi olla toiminnallinen ja että CAMP-tekijän aktiivisuudessa on kantakohtaisia eroja.
kaksi biokemiallista testijärjestelmää ei voi yksiselitteisesti erottaa F. Magnaa ”F. nericiasta”
GPAC: n biokemiallisia reaktioita, mukaan lukien Finegoldia sp: n kannat. on kuvattu; vain vähän biokemiallista vaihtelua Finegoldia sp: n välillä. kantoja raportoitiin aiemmin 2, 7. Testasimme kahta kaupallisesti saatavilla olevaa biokemiallista testijärjestelmää, joita käytetään anaerobien tunnistamiseen. Paketti ”API® 20A” sisältää 21 testiä, joista 16 testiä hiilihydraattien hyödyntämiseksi. Kuten aiemmin raportoitiin 7, mikään finegoldia sp ei voi metaboloida hiilihydraatteja. kanta (tietoja ei näytetä). Muut reaktiot (indoli, ureaasi, katalaasi, eskuliinin hydrolyysi ja gelatiinin nesteytys) eivät myöskään paljastaneet eroa F. magna-ja ”F. nericia” – kantojen välillä (tiedot eivät näy). ”RapIDTM ANA II” – pakkaus sisältää 18 entsymaattisen toiminnan testiä. Siinä missä sakkarolyyttiset entsyymit puuttuvat, kaikki Finegoldia sp. kannat tuottavat proteolyyttisiä entsyymejä (taulukko S5). Fosfataasiaktiivisuutta havainnut reaktio oli ainoa kantojen välillä poikkeava reaktio, jossa kaikki F. magna-kannat olivat ilmeisesti negatiivisia ja useimmat ”F. nericia” – kannat positiivisia. Tämän testituloksen tulkinta on kuitenkin rajallinen, sillä värinmuutos oli merkitsevyyden rajalla. Yhteenvetona voidaan todeta, että kaksi olemassa olevaa kaupallista biokemiallista testijärjestelmää, API® 20 A ja RapIDTM ANA II, eivät voi yksiselitteisesti erottaa F. Magnaa ”F. nericiasta”. Finegoldia sp: n biokemialliset ominaisuudet. niitä on tutkittava lisää biokemiallisilla lisäkokeilla.
mikroskopia paljastaa pinnan erot F. Magnan ja”F. nerician ”välillä
yrittäessään visualisoida F. Magnan ja” F. Magnan morfologiaa ja pintarakennetta. nericia ” käytimme pyyhkäisyelektronimikroskopiaa (sem) ja atomivoimamikroskopiaa (AFM) F. Magnan (07T609, 08T492 ja 09t408) ja ”F. nerician” (09T494, 12T272 ja 12t306) kantoihin. SEM havaitsi pintaeroja: F. Magnan solut olivat kiinnittyneempiä tai aggregatiivisia, näennäisesti tuottaen solunulkoisen polymeerimatriisin (Fig. 6). Tämä vahvistaa aiemman työn, joka on visualisoinut eksopolysakkaridimatriisin F. Magnan biofilmeissä (kanta FmBs12) pyyhkäisyelektronimikroskopialla sekä konfokaalisella laserkeilausmikroskopialla 23. Testatussa ”F. nerician ” kantoja emme voineet tunnistaa tällaista matriisia; solut näyttävät vähemmän aggregoituvilta.
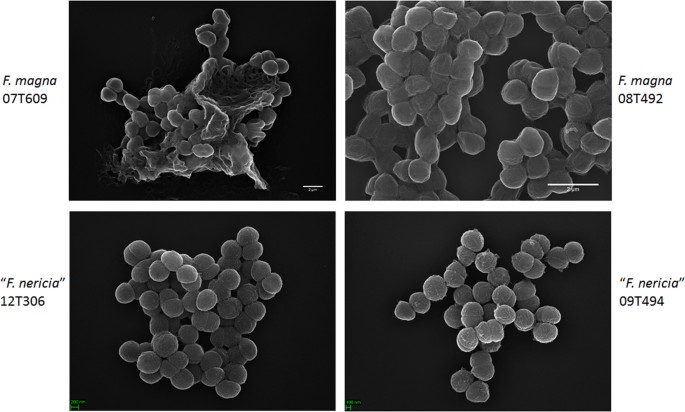
Pyyhkäisyelektronimikroskooppinen vertailu F. Magnan ja ”F. nerician”välillä. Ylä-ja alapaneeleissa on edustavia kuvia F. magna 07T609 ja 08T492 sekä ”F. nericia” 09T494 ja 12t306. F. magna-kantojen solut ovat kiinnittyneempiä/aggregatiivisia ja näyttävät tuottavan polymeerimatriisin, erityisesti kannan 07T609; sen sijaan ”F. nerician” solut ovat vähemmän aggregatiivisia.
AFM vahvisti nämä huomautukset. Lisäksi havaitsimme eroja solulisäkkeiden suhteen. F. Magnan solut tuottivat pidempiä rihmamaisia lisäkkeitä, jotka työntyivät ulos koko solun pinnasta; nämä rakenteet saattoivat esittää piliä tai fimbriae (Kuva. 7). Sen sijaan ”F. nericia” – kantojen soluilla oli sileämpi pinta ilman pitkiä ulkonevia lisäkkeitä. Ei kuitenkaan voida sulkea pois sitä, että nämä AFM-havainnot riippuvat bakteerien viljelyolosuhteista ja näytteiden käsittelyvaiheista.

atomivoiman mikroskooppivertailu F. Magnan ja ”F. nerician”välillä. Vasen ja oikea paneeli esittävät F. magna 07t609: n ja ”F. nericia” 12T306: n kuvia. (A ja C) korkearesoluutioisia kuvia avustetulla edistyneellä soft-touch-menetelmällä, jossa Z-asteikkotangot ovat enintään 0,3 µm ja XY-tangot osoittavat 500 nm. (B ja D) Overlay kuvia korkean kontrastin kuvia (a ja C), yhdistettynä riippumaton vaaka-ja vertikaalinen polynomi poisto (A ja C), ratkaistu, että 11.aste. Kokeita tehtiin kolmella näytteellä kustakin kannasta.