- genomy szczepów Finegoldia
- podział szczepów Finegoldii na dwa odrębne klady w oparciu o porównanie genomu
- heterogeniczność specyficzna dla szczepu Finegoldia sp
- czynniki oddziałujące na gospodarza i (przypuszczalne) czynniki zjadliwości
- genomy Finegoldii kodują czynniki CAMP, które są funkcjonalne
- dwa systemy badań biochemicznych nie mogą jednoznacznie odróżnić F. magna od „F. nericia”
- mikroskopia ujawnia różnice powierzchni między F. magna i „F. nericia”
genomy szczepów Finegoldia
w sumie 17 genomów Finegoldia sp. analizowano i porównywano. Dziesięć szczepów zostało niedawno zsekwencjonowanych przez naszą grupę15; zostały one wyizolowane od pacjentów z infekcjami związanymi z ortopedycznymi implantami stawów w Szwecji. Siedem genomów zostało wcześniej zsekwencjonowanych przez inne, w tym zamknięty Genom szczepu typu ATCC 2932814. Cechy genomu wszystkich dotychczas zsekwencjonowanych szczepów Finegoldii podsumowano w tabeli 1. Adnotacja z użyciem Prokka wykryła między 1570 a 1906 sekwencjami kodującymi (CDS) na Genom, ze średnią 1760 CD na szczep.
podział szczepów Finegoldii na dwa odrębne klady w oparciu o porównanie genomu
wcześniej zauważyliśmy dużą heterogeniczność między genomami izolatów Finegoldii15. Przeprowadzono wyrównanie rdzenia do genomu wszystkich 17 genomów i nazwano polimorfizmy pojedynczego nukleotydu (SNPs) w genomie rdzeniowym. W sumie, 126,647 Rdzeń genomu SNP zidentyfikowano i wykorzystano do filogenomicznej rekonstrukcji, ujawniając, że 17 genomy mogą być zgrupowane w dwa odrębne klady (Fig. 1). Dodatkowa analiza, która określa średnią identyfikację nukleotydów (ANI) genomu rdzenia wykazała, że oba klady wykazują ANI 90,7%, z odmianową zmiennością ANI od 90,2% do 91,2%. Tak niski wskaźnik ANI wskazuje, że oba klady reprezentują poszczególne gatunki16. Jeden Klad zawiera większość wcześniej zsekwencjonowanych i badanych szczepów, w tym ATCC29328 i ALB8; można go uznać za klasyczny Klad F. magna, z ani intraclade 96,6% (zmienność 95,1–97,4%). Drugi Klad z intraclade ANI 94,8% (93,4-96.6%) zawiera inny szczep ATCC (ATCC 53516) i głównie szczepy wyizolowane od pacjentów ze Szpitala Uniwersyteckiego w Örebro w Szwecji15. Następnie szczepy tego kladu są wstępnie nazywane „Finegoldia nericia” (łac. Närke, region Szwedzki, gdzie wyizolowano większość szczepów tego gatunku).

drzewo Filogenomiczne oparte na SNP genomu rdzeniowego wszystkich 17 sekwencjonowanych genomów szczepów Finegoldia. Program Pasternp został użyty do wyrównania genomu rdzenia i wywołania SNP. Genom rdzeniowy jest reprezentowany przez 43% genomu referencyjnego (ATCC29328); w sumie 126 647 wiarygodnych SNP genomu rdzeniowego wykorzystano do rekonstrukcji filogenezy całego genomu. Genomy można przypisać do dwóch głównych kladów; jeden Klad reprezentuje szczepy F. magna, a drugi Klad składa się ze szczepów nowego gatunku, tutaj wstępnie nazwanego „F. nericia”.
porównanie sekwencji 16S rRNA wszystkich izolatów wykazało wysoki stopień ogólnego podobieństwa (>99%), ale wykryło oddzielenie izolatów F. magna od izolatów „F. nericia”, ze względu na 5 do 9 „F. nericia ” – specyficzne SNP (rysunek S1). Ujawnił również większe zróżnicowanie wśród izolatów” F. nericia ” w porównaniu z F. magna. Następnie przeanalizowaliśmy dodatkowe sekwencje 16S rRNA należące do Finegoldia Sp. przechowywany w Genbanku (rysunek S2). Ujawniło to, że większość przechowywanych sekwencji klastruje się z „F. nericia”, w szczególności z grupy Finegoldia sp. szczepy wyizolowane z biofilmów ze stentu żółciowego17. Ponownie wykryto większe zróżnicowanie wśród izolatów „F. nericia”. Następnie przeanalizowaliśmy ludzkie dane mikrobiomu przechowywane w bazie danych img (Integrated Microbial Genomes and microbiome samples) pod kątem obecności sekwencji F. magna i „F. nericia”. 3287 i 7068 sekwencje kodujące F. magna ATCC29328 i” F. nericia ” ATCC 53516, odpowiednio, mogły być zidentyfikowane w 875 zebranych ludzkich zestawach danych dotyczących mikrobiomu obecnych w IMG (dane nie pokazane); najwyższe występowanie Finegoldii wykryto w okazach zgięcia wstecznego. Łącznie analizy te sugerują, że wśród spokrewnionych z człowiekiem Finegoldii izolat „F. nericia” wydaje się być bardziej rozpowszechniony niż F. magna.
heterogeniczność specyficzna dla szczepu Finegoldia sp
aby rzucić dalsze światło na strukturę populacji rodzaju Finegoldia, przeprowadzono analizy porównawcze 12 genomów (wszystkie dziesięć nowo zsekwencjonowanych szczepów i dwa szczepy ATCC), w tym cztery szczepy kladu F. magna i osiem szczepów kladu „F. nericia”. Wizualne porównanie 12 genomów podkreśliło różnicę tożsamości nukleotydów między dwoma kladami (Fig. 2). Co ciekawe, plazmid znaleziony w ATCC 29328 wydaje się być specyficzny dla tego szczepu, ponieważ żaden inny zsekwencjonowany szczep nie zawiera podobnej sekwencji (Fig. 2A).

porównanie genomu 12 szczepów należących do rodzaju Finegoldia. Dwa najbardziej wewnętrzne pierścienie reprezentują ZAWARTOŚĆ G + C (Czarny) i GC-skew (fioletowy/zielony). (A) szczepem referencyjnym jest atcc29328 (F. magna); zwróć uwagę na wysoką tożsamość nukleotydów (wizualizowaną przez ciemnoczerwony kolor) trzech najbardziej wewnętrznych genomów (należących wszystkie do F. magna), a niższy identyfikuje się z ośmioma szczepami należącymi do „F. nericia” (bladoczerwony kolor ośmiu zewnętrznych pierścieni); od wewnątrz na zewnątrz: szczepy 07t609, 08t492, 09t408, 09T494, 12t272, 12T273, 12T306, CCUG 54800, T151023, T160124 i ATCC 53516. Duży region specyficzny dla ATCC29328 w lewym górnym obszarze reprezentuje plazmid pFMC. B) szczepem referencyjnym jest 09t494 („F. nericia”). Siedem genomów „F. nericia” (pierścienie wewnętrzne) ma wyższą identyfikowalność nukleotydów (reprezentowaną przez ciemniejszy czerwony kolor), w porównaniu z czterema genomami F. magna (pierścienie zewnętrzne); od wewnątrz do zewnątrz: szczepy 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 i 09T408. Nie ma oznak specyficznych dla”F. nericia” regionów genomowych, ale zwróć uwagę na pięć dużych skupisk specyficznych dla szczepów.
nie stwierdzono istotnych różnic w wielkości genomu i liczbie CD pomiędzy tymi dwoma kladami. Ponadto nie wykryto regionów genomowych specyficznych dla kladu (tj. F. magna lub „F. nericia”), jak oceniono na podstawie analizy BRIG (Fig. 2), a także potwierdzone przez dwukierunkowe podejście blast wszystkich CDS przy użyciu ProteinOrtho (tabela S1). Podejście to wykryło 1202 ortologów współdzielonych przez wszystkie 12 genomów, tj. średnio 68% CDS każdego szczepu jest częścią proteomu rdzenia. Znaleziono tylko nieliczne geny specyficzne dla kladu. Zamiast tego można znaleźć wiele regionów dzielonych przez tylko kilka szczepów, np. dla szczepu 09t494 (patrz także Fig. 2b). Ponadto obecne są regiony specyficzne dla szczepu, które kodują w sumie 1016 specyficznych dla szczepu CDS, z dużym zakresem między szczepami, np. 187 i 22 specyficzne dla szczepu CDS wykrywa się odpowiednio w szczepie ATCC29328 i T151023 (tabela S1). Wśród funkcji specyficznych dla szczepu są różne funkcje transportowe (żelazo, pompy wypływu, transportery ABC OLIGOPEPTYDU), białka powierzchniowe, systemy modyfikacji restrykcyjnej, funkcje transferu koniugacyjnego, synteza poliketydów, bakteriocyny, funkcje związane z fagami, determinanty oporności na antybiotyki i systemy CRISPR/cas.
co ciekawe, wykryto różnice w systemach CRISPR / cas pomiędzy szczepami F. magna i „F. nericia” (tabela S2). Siedem z dziewięciu F. szczepy magna zawierały jedną do czterech macierzy CRISPR na szczep, zawierających średnio 12 odstępów (od 3 do 38 odstępów na szczep). Powtórzenie „GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC” było specyficzne dla szczepów F. magna. Natomiast szczepy „F. nericia” nosiły tylko jeden układ CRISPR na szczep, który zawierał średnio 53 przekładki (zakres od 14 do 87 na szczep). Każdy szczep zawierał indywidualny zestaw przekładek w ich tablicy CRISPR, co wskazywało na heterogeniczność specyficzną dla szczepu i wskazywało, że każdy szczep ma unikalną historię ewolucyjną.
aby ocenić, czy takie specyficzne dla szczepu funkcje zostały nabyte poziomo, przyjrzeliśmy się oznakom ich mobilności/nabycia za pomocą programu IslandViewer (rysunek S3). Program ten przewidywał 141 (szczep 09t408) do 408 (szczep 12t273) genów zlokalizowanych na wyspach genomowych, które prawdopodobnie zostaną nabyte poziomo, w tym wiele wyżej wymienionych genów specyficznych dla szczepu zaangażowanych w specjalizację, sprawność, przetrwanie i konkurencję międzygatunkową (tabela S3).
czynniki oddziałujące na gospodarza i (przypuszczalne) czynniki zjadliwości
opisano kilka czynników oddziałujących na gospodarza dla F. magna 4,5. Zidentyfikowano pili zależne od sortazy, które są ważne dla adhezji i kolonizacji13. Locus genomowy kodujący geny dla podjednostki pilusa Fmp1, przypuszczalnej podjednostki fmp2 i trzech sortaz jest zachowywany wśród wszystkich zsekwencjonowanych genomów (Fig. 3A). Istnieją specyficzne dla szczepu odmiany w górę i w dół tego locus, tj. niektóre szczepy posiadają dodatkowe geny przed fmp1 i po locus sortazy, w tym gen kodujący białko z rodziny hemolizyn III. Co ciekawe, istnieją specyficzne dla szczepu różnice dotyczące fmp1. Prawie każdy szczep posiada indywidualny wariant, a analiza filogenetyczna Fmp1 na poziomie białka nie wykazała korelacji z analizą filogenomiczną genomu rdzenia (Fig. 3b). Natomiast Fmp2 jest wysoce zachowywany wśród wszystkich szczepów. Inne białko, kodowane przed fmp1, zawiera domeny wiążące kolagen i fibrynogen; domena wiążąca kolagen (cog4932) jest również obecna w Fmp2. Proponujemy, aby białko to również należało do pilus locus i nazwało odpowiedni Gen fmp3.

pili zależne od Sortazy-kodowanie genomowych loci w genomach szczepów Finegoldia. A) Genomowa organizacja loci u szczepów F. magna (czerwony) i „F. nericia” (Jasnoniebieski). Trzy geny sortazy są przedstawione w kolorze jasnozielonym (oznaczone jako „2”); gen dla głównej podjednostki pilusa Fmp1 jest pokazany na Czerwono (oznaczony „1”), A fmp2 na pomarańczowo (oznaczony”3″). Przed fmp1 i przed genami sortazy w niektórych szczepach obecne są dodatkowe geny (oznaczane jako” 8″, białko z rodziny hemolizyn III i” 10″, białko hipotetyczne). Przypuszczalne białko wiążące kolagen, FMP3 (oznaczone jako „4”), jest kodowane przed fmp1. B) drzewo filogenetyczne fmp1 szczepów Finegoldia. Większość szczepów posiada indywidualny wariant Fmp1. Historia ewolucji została wywnioskowana za pomocą metody maksymalnego prawdopodobieństwa, wykonanej w MEGA7.
białko L jest prawdopodobnie najlepiej zbadanym czynnikiem F. magna 8. Jest to superantygen, który celuje w komórki B i immunoglobuliny. Wykryto go w około 10% dotychczas badanych izolatów Finegoldii, ale różni się wielkością i liczbą domen wiążących fragment antygenu (Fab) (domena B1, pfam02246)4,8,18. Badanie blastem badanego białka L ze szczepu 312 (locus ID: M86697, 719 aa) ujawnia kilka bardzo podobnych (tożsamość białka powyżej 50%) białek o różnych rozmiarach (603 do 1417 aa) w kilku szczepach, tj. szczepy CCUG54800, T161024, ATCC53516, 12T273 i 12t306 (rys. 4a). Wszystkie są to szczepy „F. nericia”, co wskazuje, że u tego gatunku bardziej rozpowszechniony jest superantygen limfocytów B. Wyjątkowo duży (1417 aa) białkowy czynnik l-podobny w szczepie 12t306 posiada oprócz dwóch domen B1 również dwa moduły GA znane z białek wiążących albuminy peptostreptokokowe (patrz poniżej). Wiele dodatkowych białek o niskim podobieństwie do białka L, ale nie zawierających żadnych pozornych domen B1 jest kodowanych w sekwencjonowanych przez Genom szczepach Finegoldia (tabela S4A); ponownie, istnieją odmiany specyficzne dla szczepu, a kilka szczepów, tj. 09t408, 09t494 i T151023 nie posiada takich homologów.

genomowe loci kodujące białka L i homologi FAF w szczepach Finegoldia. (A) badania Blast z charakteryzowanym białkiem L ze szczepu 312 (locus ID: M86697) przeprowadzono w sekwencjonowanych genomowo szczepach Finegoldia i pokazano homologi (w kolorze czerwonym, oznaczone „1”) o wysokim podobieństwie (>50% identyczności białka) (patrz również tabela S4A). Tylko szczepy „F. nericia”, ale nie F. szczepy magna zawierają takie homologi o różnej długości. (B) badania Blast z badanym białkiem FAF szczepu ALB8 wykazały homologi pełnej długości (na Czerwono, oznaczone „1”) w trzech szczepach F. magna (czerwony) i czterech „F. nericia” (Jasnoniebieski). Gen FAF występuje w dwóch szczepach „F. nericia” t151023 i 12t272.
stwierdzono, że Finegoldia magna wiąże się z ludzkim albuminem9. W szczepie ALB8 badano peptostreptokokowe białko wiążące albuminę (PAB); jego aktywność jest związana z przyspieszonym tempem wzrostu9. Białko PAB szczepu ALB8 (locus ID: X77864) zawiera moduły GA, domenę wiążącą albuminy; struktura modułu GA została rozwiązana19. Badanie Blast ujawniło, że w innych sekwencjonowanych genomach Finegoldia nie można znaleźć Bliskiego HOMOLOGA PAB (tabela S4B). Zamiast tego istnieje kilka homologów, które dzielą do 50% tożsamości białka; podobieństwa homologów w szczepach F. magna są średnio wyższe w porównaniu z homologami w szczepach „F. nericia”. Jeśli chodzi o białka zawierające moduł GA, w sumie 16 białek jest kodowanych w 12 genomach Finegoldii, co wskazuje na pewną redundancję białek wiążących albuminy (tabela S4C). Większość szczepów koduje dwa białka z modułami GA, z wyjątkiem szczepu 09t408 („F. magna”) i T151023 („F. nericia”).
wykazano, że subtylizynopodobna proteaza Sufa przyłączona do ściany komórkowej degraduje kilka białek ludzkich, takich jak kolagen IV,fibrynogen i peptydy przeciwdrobnoustrojowe,takie jak LL-37 I mig/CXCL910, 20, 21. Badania Blast wykazały, że większość szczepów Finegoldia posiada homolog SufA, który jest bardzo podobny (tożsamość białka > 75%) do badanego SufA ze szczepu ALB8 (locus ID: DQ679960). Wyjątkami są szczepy 07T609, 09T408 i CCUG54800. Drugi homolog SufA jest obecny w większości szczepów; średnia tożsamość białka wynosi 26% (tabela S4D).
innym ważnym czynnikiem oddziałującym na gospodarza jest FAF (F. magna adhezja factor). Około 90% szczepów F. magna wytwarza ten czynnik, który jest odpowiedzialny za zbrylanie bakterii i pośredniczy w wiązaniu z błoną podstawną poprzez wiązanie z BM-4011. Okazało się, że siedem (trzy szczepy F. magna i cztery szczepy „F. nericia”) z 12 szczepów koduje homolog FAF podobny do tego badanego w szczepie ALB8 (Fig. 4B). Istnieją różnice specyficzne dla szczepu i mutacje frameshift są widoczne u szczepów” F. nericia ” t151023 i 12t272. Nie znaleziono homologu w szczepach 09t408, 09t494 i 12t306 (tabela S4E).
genomy Finegoldii kodują czynniki CAMP, które są funkcjonalne
przeszukaliśmy Genom pod kątem innych domniemanych czynników oddziałujących na gospodarza i znaleźliśmy geny kodujące czynniki Christie-Atkins-Munch-Petersen (CAMP). Czynniki CAMP mogą działać jako kohemolizyny i odpowiadać za tak zwaną reakcję CAMP, synergiczną lizę erytrocytów owiec przez sfingomielinazę C (toksynę beta) Staphylococcus aureus i czynnik CAMP22.
eksploracja genomu zidentyfikowała dwa geny kodujące czynniki CAMP w każdym sekwencjonowanym przez Genom szczepie Finegoldia (CAMP1 i CAMP2; rysunek S4); każdy homolog zawiera domenę rodziny czynników CAMP (Pfam07373). Co ciekawe, jeden homolog (CAMP2) na szczep zawiera dodatkową bakteryjną domenę podobną do IG (Pfam02368). Oprócz CAMP1 i CAMP2, cztery ” F. szczepy „nericia” zawierają dwa dodatkowe czynniki CAMP, które znacznie różnią się od innych homologów (CAMP3 i CAMP4; rysunek S4). Tak więc cztery z siedmiu szczepów „F. nericia”posiadają cztery geny CAMP factor.
następnie chcieliśmy wiedzieć, czy szczepy Finegoldia wywołują pozytywną reakcję CAMP. Test płytki agarowej CAMP pokazuje, że niektóre szczepy są silnie dodatnie od reakcji CAMP, podczas gdy dla innych szczepów tylko niewielka kohemoliza może być wykryta w zastosowanych warunkach uprawy w dwóch i pięciu dniach inkubacji (Fig. 5, Rysunek S5). Wszystkie trzy testowane F. szczepy magna były silnie CAMP-dodatnie. Wśród szczepów” F. nericia ” wykryto pewną heterogeniczność; dwa szczepy były silnie dodatnie ODCZYNOWO CAMP, cztery szczepy były tylko nieznacznie kohemolityczne, a jeden szczep, CCUG54800, był wyraźnie ujemny odczynowo Camp. Analiza genomiczna wykazała, że w szczepie CCUG54800 gen kodujący CAMP1 zawierał wstawienie locus kodującego transpozazę w 5′-końcu genu, co zaburza Gen (rysunek S6).

reakcja CAMP szczepów Finegoldia. Zastosowano płytki agarowe TSA z 5% krwią owczą. Staphylococcus aureus jest zaszczepiony w pasie środkowym. Pozytywny wynik testu CAMP wskazuje całkowita liza erytrocytów na styku Finegoldia Sp. i smugi S. aureus. Wszystkie trzy szczepy F. magna (w kolorze czerwonym) i dwa z siedmiu szczepów „F. nericia” (w Kolorze Niebieskim) wykazały silną pozytywną reakcję CAMP po dwóch dniach beztlenowej inkubacji.
nasze wyniki wskazują, że czynniki CAMP Finegoldia Sp. może być funkcjonalny i że istnieją specyficzne dla szczepu różnice w aktywności czynnika CAMP.
dwa systemy badań biochemicznych nie mogą jednoznacznie odróżnić F. magna od „F. nericia”
biochemicznych reakcji GPAC, w tym szczepów Finegoldia Sp. zostały opisane; tylko niewielka zmienność biochemiczna wśród Finegoldia Sp. szczepy były wcześniej zgłaszane2, 7. Przetestowaliśmy dwa dostępne na rynku biochemiczne systemy testowe, które są używane do identyfikacji beztlenowców. Zestaw „API® 20A” zawiera 21 testów, w tym 16 testów wykorzystania węglowodanów. Jak donoszono poprzednio 7, żaden z węglowodanów nie może być metabolizowany przez Finegoldia Sp. szczep (brak danych). Pozostałe reakcje (indol, ureaza, katalaza, hydroliza eskuliny i upłynnianie żelatyny) również nie wykazały żadnej różnicy między szczepami „F. magna” i „F. nericia” (dane nie zostały przedstawione). Zestaw „RapIDTM ANA II” zawiera 18 testów aktywności enzymatycznej. Podczas gdy brakuje enzymów sacharolitycznych, wszystkie Finegoldia sp. szczepy wytwarzają enzymy proteolityczne (tabela S5). Reakcja wykrywająca aktywność fosfatazy była jedyną reakcją, która różniła się między szczepami, przy czym wszystkie szczepy „F. magna” okazały się ujemne, a większość szczepów” F. nericia ” dodatnie. Interpretacja tego wyniku testu jest jednak ograniczona, ponieważ zmiana koloru znajdowała się na granicy znaczenia. Podsumowując, dwa istniejące komercyjne systemy badań biochemicznych, API® 20 A i RapIDTM ANA II, nie mogą jednoznacznie odróżnić F. magna od „F. nericia”. Właściwości biochemiczne Finegoldia Sp. muszą być dalej badane z dodatkowymi testami biochemicznymi.
mikroskopia ujawnia różnice powierzchni między F. magna i „F. nericia”
w próbie wizualizacji morfologii i struktury powierzchni F. magna i ” F. nericia ” użyliśmy skaningowej mikroskopii elektronowej (SEM) i mikroskopii sił atomowych (AFM) na szczepach F. magna (07t609, 08t492 i 09t408) i „F. nericia” (09T494, 12T272 i 12T306). Sem wykryto różnice powierzchniowe: komórki F. magna były bardziej przylegające lub agregacyjne, pozornie wytwarzając pozakomórkową matrycę polimerową (Fig. 6). Potwierdza to Wcześniejsze prace, które wizualizowały matrycę egzopolisacharydów w biofilmach F. magna (szczep fmbs12) za pomocą skaningowej mikroskopii elektronowej oraz konfokalnej laserowej mikroskopii skaningowej 23. W testowanym ” F. nericia ” szczepy nie mogliśmy zidentyfikować takiej matrycy; komórki wydają się mniej agregacyjne.
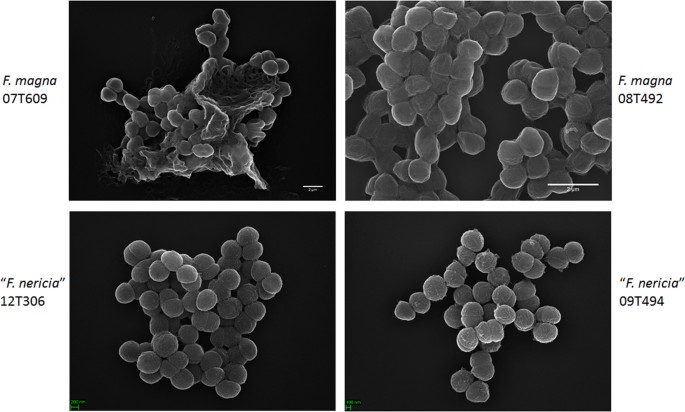
Scanning electron microscopic comparison of F. magna and „F. nericia”. Górne i dolne panele pokazują reprezentatywne obrazy F. magna 07t609 i 08t492 oraz” F. nericia ” 09t494 i 12t306, odpowiednio. Komórki szczepów F. magna są bardziej przylegające/agregatywne i wydają się wytwarzać matrycę polimerową, w szczególności szczep 07t609; natomiast komórki „F. nericia” są mniej agregatywne.
AFM potwierdziło te obserwacje. Ponadto zaobserwowaliśmy różnice dotyczące przydatków komórkowych. Komórki F. magna wytwarzały dłuższe nitkowate wyrostki wystające z całej powierzchni komórki; struktury te mogły reprezentować pili lub fimbriae (Fig. 7). Natomiast komórki szczepów „F. nericia” miały gładszą powierzchnię bez długich wystających wyrostków. Nie można jednak wykluczyć, że obserwacje AFM zależą od warunków uprawy bakterii i etapów przetwarzania próbki.

Atomic force microscopic comparison of F. magna and „F. nericia”. Lewy i prawy panel przedstawiają odpowiednio obrazy F. magna 07t609 i” F. nericia ” 12t306. (A i C) obrazy o wysokiej rozdzielczości ze wspomaganą zaawansowaną metodą soft-touch z paskami w skali Z do 0,3 µm i paskami XY wskazującymi 500 nm. (B I D) nakładanie obrazów o wysokim kontraście obrazów (a i C), połączone z niezależnym poziomym i pionowym usuwaniem wielomianów (A I C), rozwiązane do 11 stopnia. Eksperymenty przeprowadzono na trzech próbkach z każdego szczepu.