- Genomů kmenů Finegoldia
- rozdělení kmenů Finegoldia do dvou odlišných kmenů na základě srovnání genomu
- Kmen specifická heterogenita Finegoldia sp
- faktory ovlivňující hostitele a (domnělé) faktory virulence
- Finegoldia genomy kódují TÁBOR faktory, které jsou funkční
- dva systémy biochemických testů nemohou jednoznačně odlišit F. magna od „F. nericia“
- mikroskopie odhaluje povrchové rozdíly mezi F. magnou a“F. nericiou“
celkem 17 genomů Finegoldia sp. byly analyzovány a porovnány. Deset kmenů nedávno byly sekvenovány pomocí našeho group15; byly izolovány od pacientů s ortopedickou kloubu implantát, infekce spojené ve Švédsku. Sedm genomů bylo dříve sekvenováno jinými, včetně uzavřeného genomu typového kmene ATCC 2932814. Genomové rysy všech dosud sekvenovaných kmenů Finegoldia jsou shrnuty v tabulce 1. Anotace pomocí Prokky detekovala mezi 1570 a 1906 kódovacími sekvencemi (CDS) na genom, s průměrem 1760 CDS na kmen.
dříve jsme zaznamenali velkou heterogenitu mezi genomy izolátů Finegoldia15. Bylo provedeno zarovnání jádra a genomu všech 17 genomů a byly volány jednonukleotidové polymorfismy (SNP) v jádrovém genomu. Celkem 126,647 core-Modifikace genomu byly identifikovány a použity pro phylogenomic rekonstrukce, která odhalila, že 17 genomy mohou být rozděleny do dvou odlišných skupin (Obr. 1). Další analýza, která určuje průměrné nukleotidové identifikovat (ANI) základní genomu ukázala, že dvě clades vykazují ANI o 90,7%, s kmenem-zvláštní ANI odchylka od 90.2% na 91,2%. Takové nízké ANI naznačuje, že tyto dva kmeny představují jednotlivé druhy16. Jeden klad obsahuje většinu dříve sekvenovány a studoval kmeny, včetně ATCC29328 a ALB8; to může být považováno za klasické F. magna clade, s intraclade ANI z 96.6% (variace 95.1–97.4%). Druhý clade s intraclade ANI 94,8% (93,4-96.6%) obsahuje další kmen ATCC (ATCC 53516)a hlavně kmeny izolované od pacientů ve Fakultní nemocnici Örebro, Sweden15. Dále jsou kmeny tohoto kladu předběžně pojmenovány „Finegoldia nericia“ (latinsky Närke, Švédská oblast, kde byla izolována většina kmenů tohoto druhu).

Phylogenomic strom na základě core-Modifikace genomu všech 17 sekvenovaných genomů Finegoldia kmenů. Program Parsnp byl použit k zarovnání genomu jádra a volání SNP. Jádro genomu je reprezentováno 43% referenčního genomu (ATCC29328); celkem 126 647 spolehlivých SNP jádra genomu bylo použito k rekonstrukci fylogeneze celého genomu. Genomy lze přiřadit ke dvěma hlavním klad; jeden klad představuje kmeny F. magna a druhý klad se skládá z kmenů nového druhu, zde předběžně pojmenovaných „F. nericia“.
srovnání 16S rRNA sekvencí všech izolátů odhalilo vysoký stupeň celkové podobnosti (>99%), ale zjistil oddělení F. magna izoluje od „F. nericia“ izoláty, vzhledem k 5 až 9 „F. nericia “ – specifické SNP (obrázek S1). Rovněž odhalila vyšší diverzifikaci mezi izoláty „F. nericia“ ve srovnání s F.magnou. Dále jsme analyzovali další 16S rRNA sekvence patřící do Finegoldia sp. uloženo v GenBank (obrázek S2). To odhalilo, že většina uložených sekvencí se shlukuje s „F. nericia“, zejména skupinou Finegoldia sp. kmeny izolované z biofilmů žlučových stentů17. Opět je zjištěna vyšší diverzifikace mezi izoláty „F. nericia“. Dále jsme analyzovali lidského mikrobiomu data uložená v IMG (Integrované Mikrobiální Genomy a mikrobiomu vzorky) databáze na přítomnost F. magna a „F. nericia“ sekvence. 3287 a 7068 kódující sekvence F. magna ATCC29328 a „F. nericia“ ATCC 53516, respektive, může být identifikován v 875 sestaven lidského mikrobiomu datových souborů přítomných v IMG (údaje nejsou uvedeny); nejvyšší výskyty Finegoldia byly zjištěny v retroaurikulární rýhy exempláře. Dohromady tyto analýzy naznačují, že mezi izoláty Finegoldia spojenými s člověkem se zdá, že“ F.nericia “ je častější než F. magna.
Kmen specifická heterogenita Finegoldia sp
vrhnout další světlo na strukturu populace Finegoldia rodu, srovnávací analýzy 12 genomů (všech deset nově sekvenovaných kmenů a dva ATCC kmenů) byly provedeny, včetně čtyř kmenů F. magna clade a osm kmenů „F. nericia“ clade. Vizuální srovnání 12 genomů zdůraznilo rozdíl nukleotidové identity mezi těmito dvěma klad (obr. 2). Je zajímavé, že plazmid nalezený v ATCC 29328 se zdá být specifický pro tento kmen, protože žádný jiný sekvenovaný kmen neobsahuje podobnou sekvenci (obr. 2A).

srovnání Genomu z 12 kmenů patřících do rodu Finegoldia. Dva nejvnitřnější kroužky představují obsah G + C (černý) a GC-zkosení (Fialová / Zelená). (A) referenční kmen je ATCC29328 (F. magna); všimněte si vysoké nukleotidové identity (vizualizované tmavě červenou barvou) tří nejvnitřnějších genomů (patřících všem F. magna), a nižší identifikovat až osm kmenů, které patří k „F. nericia“ (světle červené barvy z osmi vnější kroužky); zevnitř ven: kmeny 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 a ATCC 53516. Velká oblast specifická pro ATCC29328 v levé horní oblasti představuje plazmid pFMC. B) referenční kmen je 09T494 („F. nericia“). Sedm genomů“ F. nericia “ (vnitřní kruhy) má vyšší nukleotid (reprezentovaný tmavší červenou barvou), ve srovnání se čtyřmi genomy F. magna (vnější kruhy); zevnitř ven: kmeny 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 a 09T408. Neexistují žádné známky genomických oblastí specifických pro „F. nericia“, ale všimněte si pěti velkých kmenově specifických shluků.
Žádné významné rozdíly ve velikosti genomu a počtu DISKŮ byla zjištěna mezi dvěma clades. F. magna nebo“F. nericia“ specifické) genomové oblasti, jak bylo posouzeno z analýzy BRIG(obr. 2) a také potvrzeno obousměrným blastovým přístupem všech CDS pomocí ProteinOrtho (tabulka S1). Tento přístup detekoval 1202 ortologů sdílených všemi 12 genomy, tj. v průměru 68% CDS každého kmene je součástí jádra proteomu. Nachází se jen velmi málo genů specifických pro clade. Místo toho lze nalézt mnoho oblastí sdílených pouze několika kmeny, např. pro kmen 09T494 (viz také obr. 2B). Kromě toho, napětí-specifické oblasti jsou přítomny, které kódují celkem 1016 kmen-konkrétní CD, s velkým rozsahem mezi kmeny, např. 187 a 22 kmen-konkrétní CD jsou detekovány v napětí ATCC29328 a T151023, respektive (Tabulky S1). Mezi kmen-specifické funkce jsou různé dopravní funkce (železo, efluxní pumpy, oligopeptid ABC transportéry), povrchové proteiny, omezení-modifikace systémů, conjugative přenosové funkce, polyketide syntéza, bakteriociny, phage-související funkce, determinanty rezistence k antibiotikům, a CRISPR/cas systémy.
zajímavé je, že byly zjištěny rozdíly v systémech CRISPR/cas mezi kmeny F. magna a „F. nericia“ (tabulka S2). Sedm z devíti F. kmeny magna obsahovaly jedno až čtyři pole CRISPR na kmen, obsahující v průměru 12 rozpěrek (v rozmezí od 3 do 38 rozpěrek na kmen). Opakování „GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC“ bylo specifické pro kmeny F.magna. Naproti tomu kmeny „F. nericia“ nesly pouze jedno pole CRISPR na kmen, které obsahovalo v průměru 53 rozpěrek(rozmezí 14 až 87 na kmen). Každý kmen obsahoval individuální sadu distančních v jejich CRISPR pole(y), podřízená kmen specifická heterogenita, a naznačuje, že každý kmen má jedinečnou evoluční historii.
vyhodnotí se, zda takový kmen-specifické funkce byly horizontálně získané podívali jsme se na znamení jejich mobility/akvizice pomocí IslandViewer programu (Obrázek S3). Tento program předpověděl 141 (kmen 09T408) 408 (kmen 12T273) geny nacházející se v genomických ostrovů, které jsou pravděpodobné, že bude horizontálně získané, včetně mnoha z výše uvedených kmen-specifických genů zapojených v zaměření, fitness, přežití a mezidruhové konkurence (Tabulka S3).
faktory ovlivňující hostitele a (domnělé) faktory virulence
pro F. Magnu 4,5 bylo popsáno několik faktorů ovlivňujících hostitele. Byly identifikovány pili závislé na Sortase, které jsou důležité pro adhezi a kolonizaci13. O genomické lokus, který kóduje geny pro pilus podjednotky Fmp1, domnělý podjednotky Fmp2 a tři sortases je konzervováno u všech sekvenovaných genomů (Obr. 3A). Existují variace specifické pro kmen nahoru a po proudu od tohoto lokusu, tj. některé kmeny mají další geny před fmp1 a za sortase locus, včetně gen kódující hemolyzinový III rodinu bílkovin. Zajímavé je, že existují rozdíly specifické pro kmen týkající se fmp1. Téměř každý kmen nese individuální varianta a fylogenetická analýza Fmp1 na proteinové úrovni ukázal žádnou souvislost s phylogenomic analýza core genomu (Obr. 3B). Naproti tomu Fmp2 je vysoce konzervovaný mezi všemi kmeny. Další protein, kódovaný před fmp1, skrývá kolagen – a fibrinogen vazebné domény; doména vázající kolagen (COG4932) je také přítomna ve Fmp2. Navrhujeme, aby tento protein také patřil k lokusu pilus a pojmenoval odpovídající Gen fmp3.

Sortase závislé pili-kódování genomových lokusů v genomu Finegoldia kmenů. A) genomická organizace lokusů u kmenů F.magna (červená) a „F. nericia“ (Světle modrá). Tři geny sortázy jsou zobrazeny světle zeleně (označené „2“); gen pro hlavní podjednotku pilus Fmp1 je zobrazen červeně (označeno „1“) a fmp2 je zobrazen oranžově(označeno „3“). Před fmp1 a za sortase geny v některé kmeny další geny (označené „8“, hemolyzinový III-family protein a „10“, hypotetický protein) jsou přítomny. Domnělý protein vázající kolagen, Fmp3 (označený „4“), je kódován před fmp1. B) fylogenetický strom Fmp1 kmenů Finegoldia. Většina kmenů má individuální variantu Fmp1. Evoluční historie byla odvozena pomocí metody maximální pravděpodobnosti, provedené v MEGA7.
Protein L je pravděpodobně nejlépe studovaným faktorem F. magna 8. Je to superantigen, který se zaměřuje na B buňky a imunoglobuliny. Byl detekován u asi 10% dosud testovaných izolátů Finegoldia, ale liší se velikostí a počtem domén vázajících fragment vázající antigen (Fab) (doména B1, pfam02246)4,8,18. Výbuch vyhledávání studoval protein L z kmene 312 (locus ID: M86697, 719 aa) odhaluje několik velmi podobné (protein identity nad 50%) proteiny s různou velikostí (603 k 1417 aa) v několika kmenů, tj. kmeny CCUG54800, T161024, ATCC53516, 12T273 a 12T306 (obr. 4A). To vše jsou kmeny „F. nericia“, což naznačuje, že superantigen B-buněk je u tohoto druhu častější. Mimořádně velký (1417 aa) protein, L-jako faktor v napětí 12T306 má kromě dvou B1 domény také dva GA moduly, které jsou známé z peptostreptococcal albumin-binding proteiny (viz níže). Mnoho dalších proteinů s nízkou podobnost bílkovin L, ale bez obsahujících jakékoli zjevné B1 domény jsou kódovány v genomu-sekvenován Finegoldia kmenů (Tabulka S4A); opět existují variace specifické pro kmen a několik kmenů, tj. 09T408, 09T494 a T151023, nemá takové homology.

Genomové lokusy kódující protein L a FAF homologů v Finegoldia kmenů. (A) Výbuch hledání se vyznačuje bílkovin L z kmene 312 (locus ID: M86697) byla provedena v genomu sekvence Finegoldia kmenů, a jeho homologů (v červené, označené „1“) s vysokou podobnost (>50% bílkovin identity) jsou uvedeny (viz také Tabulka S4A). Pouze kmeny“ F. nericia“, ale ne F. kmeny magna obsahují takové homology s různou délkou. (B) Blast vyhledávání, s studoval FAF protein kmen ALB8 našel full-délka homologů (v červené, označené „1“) ve třech F. magna (červený) a čtyři „F. nericia“ (světle modrá) kmenů. Gen FAF je frameshifted ve dvou kmenech“ F. nericia “ T151023 a 12T272.
bylo zjištěno, že Finegoldia magna se váže na lidský albumin9. V kmeni ALB8 byl studován peptostreptokokový protein vázající albumin (PAB); jeho aktivita je spojena se zrychlenou rychlostí růstu9. Protein PAB kmene ALB8 (locus ID: X77864) obsahuje moduly GA, doménu vázající albumin; struktura modulu GA byla vyřešena19. Výbuchové vyhledávání odhalilo, že blízký homolog PAB nelze nalézt v jiných sekvenovaných genomech Finegoldia (tabulka S4B). Místo toho existuje několik homologů, kteří sdílejí až 50% proteinové identity; podobnosti homologů u kmenů F. magna jsou v průměru vyšší ve srovnání s homology u kmenů“ F.nericia“. Pokud jde GA modul obsahující proteiny, celkem 16 bílkoviny jsou kódovány v 12 Finegoldia genomů, což naznačuje, některé redundance albumin-binding proteinů (Tabulka S4C). Většina kmenů kóduje dva proteiny s moduly GA, kromě kmenů 09T408 (F. magna) a T151023 („F. nericia“).
buněčné stěny připojené subtilisin-like proteázové SufA bylo prokázáno, že snížit několik lidských proteinů, jako je kolagen IV, fibrinogenu a antimikrobiální peptidy jako LL-37 a MIG/CXCL910,20,21. Výbuch hledání zjištěno, že většina Finegoldia kmeny nesou SufA homolog, který je velmi podobný (protein identity > 75%) studoval SufA z kmene ALB8 (locus ID: DQ679960). Výjimkou jsou kmeny 07T609, 09T408 a CCUG54800. Druhý Sufa homolog je přítomen ve většině kmenů; má průměrnou proteinovou identitu 26% (tabulka S4D).
dalším důležitým faktorem ovlivňujícím hostitele je FAF (F. magna adhezní faktor). Přibližně 90% kmenů F. magna produkuje tento faktor, který je zodpovědný za shlukování bakterií a zprostředkovává vazbu na bazální membránu vazbou na BM-4011. Zjistili jsme, že sedm (tři F. magna a čtyři „F. nericia“ kmenů) z 12 kmenů kódovat FAF homolog podobné studoval v ALB8 napětí (Obr. 4B). Existují rozdíly specifické pro kmen a mutace frameshiftu jsou pozorovány u kmenů“ F. nericia “ t151023 a 12T272. U kmenů 09T408, 09T494 a 12T306 (tabulka S4E) nebyl nalezen žádný homolog.
Finegoldia genomy kódují TÁBOR faktory, které jsou funkční
Jsme prohledali genom pro jiné domnělé host-interakce faktorů a nalézt geny kódující Christie-Atkins-Munch-Peterson (CAMP) faktory. Campové reakce, synergickou lýzu ovčích erytrocytů pomocí Staphylococcus aureus sfingomyelinázy C (beta-toxin) a CAMP faktor22.
Genomu těžební identifikovány dva geny kódující TÁBOR faktory v každém genomu-sekvenován Finegoldia kmen (CAMP1 a CAMP2; Obrázek S4); každý homolog přístavy TÁBOR faktor rodina domény (Pfam07373). Je zajímavé, že jeden homolog (CAMP2) na kmen obsahuje další bakteriální doménu podobnou ig (Pfam02368). Kromě CAMP1 a CAMP2 čtyři “ F. kmeny nericia “ obsahují dva další faktory CAMP, které se podstatně liší od ostatních homologů (CAMP3 a CAMP4; obrázek S4). Čtyři ze sedmi kmenů „F. nericia“ tedy obsahují čtyři geny CAMP faktoru.
dále jsme chtěli vědět, zda kmeny Finegoldia produkují pozitivní CAMP reakci. TÁBOR agar plate testu ukazují, že některé kmeny jsou silně TÁBORA reakce-pozitivní, zatímco pro jiné kmeny jen málo co-hemolýza může být detekován pod použita kultivační podmínky ve dvou a pěti dnech inkubace (Obr. 5, Obrázek S5). Všechny tři testované F. magna kmeny byly silně CAMP reakce pozitivní. Mezi „F. nericia“ kmenů jsme detekovali heterogenita; dva kmeny byly silně TÁBORA reakce-pozitivní, čtyři kmeny byly jen mírně co-hemolytická, a jeden kmen, CCUG54800, byl jasně TÁBORA reakce-negativní. Genomová analýza zjistila, že v kmen CCUG54800 v CAMP1-encoding gene obsahoval vložení transposase-kódování locus v 5′-konci genu, který narušuje gen (Obrázek S6).

TÁBOR reakce Finegoldia kmenů. Byly použity agarové destičky TSA s 5% ovčí krve. Staphylococcus aureus je inokulován ve středním pruhu. Pozitivní CAMP test je indikován úplnou lýzou erytrocytů na rozhraní Finegoldia sp. a pruhy s.aureus. Všechny tři kmeny F. magna (v červené barvě) a dva ze sedmi kmenů „F.nericia“ (v modré barvě) vykazovaly silnou pozitivní CAMP reakci po dvou dnech anaerobní inkubace.
naše výsledky ukazují, že CAMP faktory Finegoldia sp. může být funkční a že existují rozdíly specifické pro kmen v aktivitě CAMP faktoru.
dva systémy biochemických testů nemohou jednoznačně odlišit F. magna od „F. nericia“
biochemické reakce GPAC včetně kmenů Finegoldia sp. byly popsány; pouze malá biochemická variace mezi Finegoldia sp. kmeny byly dříve hlášeny2, 7. Testovali jsme dva komerčně dostupné biochemické testovací systémy, které se používají k identifikaci anaerob. Sada „API® 20A“ obsahuje 21 testů, mezi nimi 16 testů na využití sacharidů. Jak bylo uvedeno dříve7, žádný z uhlohydrátů nemohl být metabolizován žádným Finegoldia sp. kmen (údaje nejsou zobrazeny). Další reakce (indol, ureáza, kataláza, eskulin hydrolýze a zkapalnění želatiny) rovněž neodhalilo žádný rozdíl mezi kmeny F. magna a „F. nericia“ (údaje nejsou uvedeny). Souprava „RapIDTM ANA II“ obsahuje 18 testů enzymatických aktivit. Zatímco sacharolytické enzymy chybí, všechny Finegoldia sp. kmeny produkují proteolytické enzymy (tabulka S5). Reakce detekující aktivitu fosfatázy byla jedinou reakcí, která se mezi kmeny lišila, přičemž všechny kmeny F. magna byly zjevně negativní a většina kmenů“ F.nericia “ pozitivní. Interpretace tohoto výsledku testu je však omezená, protože změna barvy byla na hranici významu. Závěrem lze říci, že dva existující komerční biochemické testovací systémy, API® 20 A a RapIDTM ANA II, nemohou jednoznačně odlišit F. Magnu od „F. nericia“. Biochemické vlastnosti Finegoldia sp. je třeba dále prozkoumat další biochemické testy.
mikroskopie odhaluje povrchové rozdíly mezi F. magnou a“F. nericiou“
ve snaze vizualizovat morfologii a povrchovou strukturu F. Magny a “ F. nericia“ jsme použili skenovací elektronové mikroskopie (SEM) a mikroskopie atomárních sil (AFM) na kmeny F. magna (07T609, 08T492, a 09T408) a „F. nericia“ (09T494, 12T272 a 12T306). SEM detekoval povrchové rozdíly: buňky F. magna byly více přilnavé nebo agregační, zdánlivě produkující extracelulární polymerní matrici (obr. 6). To potvrzuje dřívější práce, která se zobrazil na exopolysaccharide matice v biofilmy F. magna (kmen FmBs12) pomocí skenovací elektronové mikroskopie, stejně jako konfokální laserové skenovací microscopy23. V testovaném „F. nericia “ kmeny takovou matrici jsme nemohli identifikovat; buňky se zdají méně agregativní.
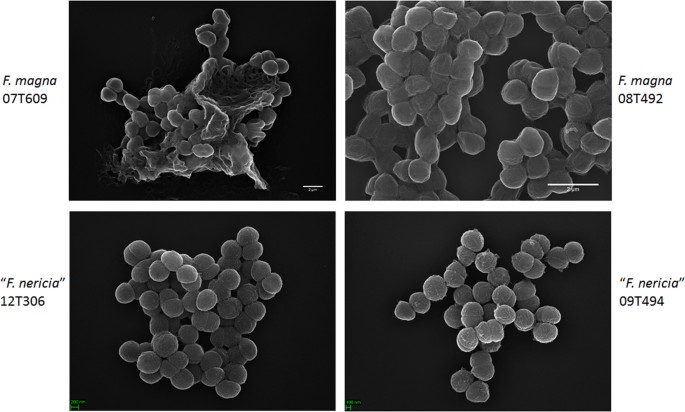
Skenování elektronovým mikroskopem srovnání F. magna a „F. nericia“. Horní a dolní panely zobrazují reprezentativní obrazy F. magna 07T609 a 08T492 a“ F.nericia “ 09T494 a 12T306. Buněk F. magna kmeny jsou více přilnavé/souhrnné a zdá se, že produkovat polymerní matrice, zejména kmen 07T609; naproti tomu, buňky „F. nericia“ jsou méně souhrnné.
AFM potvrdila tato pozorování. Kromě toho jsme pozorovali rozdíly týkající se buněčných příloh. Buňky F. Magny produkovaly delší vláknité přívěsky, které vyčnívaly z celého buněčného povrchu; tyto struktury mohly představovat pili nebo fimbrie (obr. 7). Naproti tomu buňky kmenů „F. nericia“ měly hladší povrch bez dlouhých vyčnívajících přídavků. Nelze však vyloučit, že tato pozorování AFM závisí na podmínkách kultivace bakterií a na krocích zpracování vzorku.

Atomic force mikroskopické srovnání F. magna a „F. nericia“. Levý a pravý panel představují obrazy F. Magny 07T609 a“ F.nericia “ 12T306. (A A C) obrázky s vysokým rozlišením s asistovanou pokročilou metodou soft-touch s pruhy stupnice Z až do 0,3 µm a pruhy XY označující 500 nm. (B A D) překryvné obrazy vysoce kontrastních obrazů (A A C), v kombinaci s nezávislým horizontálním a vertikálním odstraněním polynomu (A A C), vyřešeno na 11. stupeň. Pokusy byly provedeny na třech vzorcích z každého kmene.