- a Finegoldia törzsek Genomjai
- a Finegoldia törzsek felosztása két különálló kládra a genom összehasonlítása alapján
- a Finegoldia sp Törzsspecifikus heterogenitása
- gazdaszervezet-kölcsönható faktorok és (feltételezett) virulencia faktorok
- a Finegoldia genomok funkcionális CAMP faktorokat kódolnak
- két biokémiai tesztrendszer nem képes egyértelműen megkülönböztetni az F. Magnát az “F. nericia”
- a mikroszkópia az F. magna és az “F. nericia”
a Finegoldia törzsek Genomjai
összesen 17 finegoldia sp genomot. elemezték és összehasonlították. Tíz törzset nemrégiben szekvenált a csoportunk15; ortopédiai ízületi implantátummal összefüggő fertőzésekben szenvedő betegektől izolálták őket Svédországban. Korábban hét genomot szekvenáltak mások, köztük az ATCC 2932814 típusú törzs zárt genomját. Az összes eddig szekvenált Finegoldia törzs Genom jellemzőit az 1. táblázat foglalja össze. A prokka-t használó annotáció genomonként 1570 és 1906 között mutatott ki kódoló szekvenciákat (CDS), törzsenként átlagosan 1760 CDS-t.
a Finegoldia törzsek felosztása két különálló kládra a genom összehasonlítása alapján
korábban nagy heterogenitást észleltünk a Finegoldia izolátumok genomjai között15. Mind a 17 Genom mag-Genom összehangolását elvégezték, és a maggenomban egyetlen nukleotid polimorfizmust (SNP) neveztek el. Összesen 126 647 Maggenom SNP-t azonosítottak és használtak a filogenomikai rekonstrukcióhoz, feltárva, hogy a 17 Genom két különálló kládba csoportosítható (ábra. 1). Egy további elemzés, amely meghatározza a maggenom átlagos nukleotid-azonosítását (ANI), azt mutatta, hogy a két klád 90,7% – os Ani-t mutat, törzsspecifikus Ani-variációval 90,2% – ról 91,2% – ra. Az ilyen alacsony ANI azt jelzi, hogy a két klád egyedi fajokat képvisel16. Az egyik klád tartalmazza a legtöbb korábban szekvenált és vizsgált törzset, beleértve az ATCC29328–at és az ALB8-at; a klasszikus F. magna kládnak tekinthető, 96,6% – os intraclade ANI-val (95,1-97,4% variáció). A másik klád 94,8% – os intraclade ANI-val (93,4-96.6%) tartalmaz egy másik ATCC-törzset (ATCC 53516), főleg a svédországi Universrebro Egyetemi Kórház betegeiből izolált törzseket15. A továbbiakban ennek a kládnak a törzseit kísérleti jelleggel “Finegoldia nericia” – nak nevezik (latinul az N Inconkrke, a svéd régió, ahol ennek a fajnak a legtöbb törzsét izolálták).

Filogenomikus fa az összes mag-Genom SNP-je alapján 17 a Finegoldia törzsek szekvenált genomjai. A Parsnp programot a maggenom összehangolására és az SNP-k hívására használták. A maggenomot a referencia Genom 43%-A képviseli (ATCC29328); összesen 126 647 megbízható maggenom SNP-t használtunk a teljes genom filogenitásának rekonstruálására. A genomok két fő kládhoz rendelhetők; az egyik klád az F. magna törzseket képviseli, a másik klád pedig egy új faj törzseiből áll, itt kísérleti jelleggel “F. nericia”néven.
az összes izolátum 16S rRNS szekvenciájának összehasonlítása nagyfokú általános hasonlóságot mutatott (>99%), de az F. magna izolátumok elválasztását észlelte az “F. nericia” izolátumoktól, 5-9 “F. nericia ” – specifikus SNP-k (S1 ábra). Azt is feltárta, hogy az “F. nericia” izolátumok nagyobb diverzifikációt mutatnak az F. magna-hoz képest. Ezután további 16S rRNS szekvenciákat elemeztünk, amelyek a Finegoldia sp-hez tartoznak. genbankban tárolva (S2 ábra). Ez feltárta, hogy a legtöbb tárolt szekvencia az “F. nericia” – val csoportosul, különösen a Finegoldia sp. epeúti stent biofilmekből izolált törzsek17. Ismét nagyobb diverzifikációt észlelnek az “F. nericia” izolátumok között. Ezután elemeztük az IMG (Integrated Microbial Genomes and microbiome samples) adatbázisban tárolt humán mikrobiom adatokat az F. magna és az “F. nericia” szekvenciák jelenlétére. Az F. magna ATCC29328 és az “F. nericia” ATCC 53516 kódoló szekvenciáit az IMG-ben található 875 összeállított humán mikrobiom adatkészletben lehetett azonosítani (az adatok nincsenek feltüntetve); a Finegoldia legmagasabb előfordulását retroauricularis gyűrődési mintákban mutatták ki. Ezek az elemzések együttesen azt sugallják, hogy az emberhez kapcsolódó Finegoldia izolátumok között az “F. nericia” gyakoribbnak tűnik, mint az F. magna.
a Finegoldia sp Törzsspecifikus heterogenitása
a Finegoldia nemzetség populációs szerkezetének további megvilágítására 12 Genom (mind a tíz újonnan szekvenált törzs és a két ATCC törzs) összehasonlító elemzését végezték el, beleértve az F. magna klád négy törzsét és az “F. nericia” klád nyolc törzsét. A 12 Genom vizuális összehasonlítása rávilágított a két klád közötti nukleotid identitás különbségre (ábra. 2). Érdekes módon az ATCC 29328-ban található plazmid specifikusnak tűnik erre a törzsre, mivel egyetlen más szekvenált törzs sem tartalmaz hasonló szekvenciát (ábra. 2A).

a Finegoldia nemzetségbe tartozó 12 törzs Genom összehasonlítása. A két legbelső gyűrű a G + C-tartalom (fekete) és a GC-ferdeség (ibolya/zöld). (A) a referencia törzs ATCC29328 (F. magna); vegye figyelembe a három legbelső Genom magas nukleotid-azonosságát (amelyet a sötétvörös szín vizualizál) (amelyek mind F-hez tartoznak. magna), és az alsó az “F. nericia” (a nyolc külső gyűrű halványpiros színe) nyolc törzsét azonosítja; belülről kifelé: 07t609, 08t492, 09T408, 09T494, 12t272, 12t273, 12t306, CCUG 54800, T151023, T160124 és ATCC 53516 törzsek. A nagy ATCC29328-specifikus régió a bal felső régióban a pfmc plazmidot képviseli. B) a referenciatörzs 09T494 (“F. nericia”). A hét” F. nericia ” Genom (belső gyűrűk) magasabb nukleotid-azonosítóval rendelkezik (amelyet a sötétebb vörös szín képvisel), összehasonlítva a négy F. magna genommal (külső gyűrűk); belülről kifelé: a 12t272, ATCC 53516, 12T306, 12t273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08t492 és 09T408 törzsek. Nincsenek jelei az “F. nericia” -specifikus genomiális régióknak, de vegye figyelembe az öt nagy törzsspecifikus klasztert.
a két klád között nem volt szignifikáns különbség a GENOMMÉRETBEN és a CD-k számában. Ezenkívül nem mutattak ki klád-specifikus (azaz F. magna vagy “F. nericia”-specifikus) genomiális régiókat, amint azt a BRIG elemzés alapján megítélték (ábra. 2), valamint megerősítette a kétirányú robbanás megközelítés az összes CDS segítségével ProteinOrtho (táblázat S1). Ez a megközelítés 1202 ortológust észlelt mind a 12 genomban, azaz az egyes törzsek CDS-jének átlagosan 68% – a A mag proteom része. Csak nagyon kevés klád-specifikus gén található. Ehelyett sok olyan régió található, amelyen csak néhány törzs osztozik, például a 09t494 törzs esetében (lásd még az ábrát. 2B). Ezenkívül törzsspecifikus régiók vannak jelen, amelyek összesen 1016 törzsspecifikus CDS-t kódolnak, a törzsek közötti nagy tartományban, például 187, illetve 22 törzsspecifikus CDS-t mutatnak ki az ATCC29328, illetve a T151023 törzsben (S1 táblázat). A törzsspecifikus funkciók közé tartoznak a különböző transzportfunkciók (vas, efflux szivattyúk, oligopeptid ABC transzporterek), felszíni fehérjék, restrikciós módosító rendszerek, konjugatív transzferfunkciók, poliketid szintézis, bakteriocinok, fághoz kapcsolódó funkciók, antibiotikum-rezisztencia determinánsok és CRISPR/cas rendszerek.
érdekes módon a CRISPR/cas rendszerekben különbségeket mutattak ki az F. magna és az “F. nericia” törzsek között (S2 táblázat). Kilencből hét F. a magna törzsek törzsenként egy-négy CRISPR tömböt tartalmaztak, amelyek átlagosan 12 távtartót tartalmaztak (törzsenként 3-38 távtartót tartalmaztak). A “GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC” ismétlés az F. magna törzsekre jellemző volt. Ezzel szemben az” F. nericia ” törzsek törzsenként csak egy CRISPR tömböt hordoztak, amely átlagosan 53 távtartóban volt (törzsenként 14-87 tartomány). Mindegyik törzs tartalmazott egy egyedi távtartókészletet a CRISPR tömb(ek) ben, a törzsspecifikus heterogenitás alá helyezve, jelezve, hogy minden törzsnek egyedi evolúciós története van.
annak értékelése érdekében, hogy az ilyen törzs-specifikus funkciókat vízszintesen szerezték-e meg, az islandviewer program segítségével megvizsgáltuk mobilitásuk/megszerzésük jeleit (S3 ábra). Ez a program 141 (09t408 törzs) és 408 (12t273 törzs) géneket jósolt a genomi szigeteken, amelyek valószínűleg vízszintesen megszerezhetők, beleértve a fent említett törzsspecifikus géneket, amelyek részt vesznek a specializációban, a fitneszben, a túlélésben és a fajok közötti versenyben (S3 táblázat).
gazdaszervezet-kölcsönható faktorok és (feltételezett) virulencia faktorok
számos gazdaszervezet-kölcsönható faktort írtak le az F. magna 4,5 esetében. Sortáz-függő pili-ket azonosítottak, amelyek fontosak az adhézió és a kolonizáció szempontjából13. Az fmp1 pilus alegység, a feltételezett Fmp2 alegység és a három sortáz génjeit kódoló genomi lokusz konzerválódik az összes szekvenált Genom között (ábra. 3A). Ezen a lókuszon felfelé és lefelé is vannak törzsspecifikus eltérések, pl. egyes törzsek további génekkel rendelkeznek az fmp1 előtt és a sortáz lókusz után, beleértve egy hemolizin III család fehérjét kódoló gént. Érdekes módon vannak törzsspecifikus különbségek az fmp1 tekintetében. Szinte minden törzs egyedi variánst hordoz, és az fmp1 fehérje szintű filogenetikai elemzése nem mutatott összefüggést a maggenom filogenomikai elemzésével (ábra. 3B). Ezzel szemben az Fmp2 erősen konzervált az összes törzs között. Egy másik, az fmp1 előtt kódolt fehérje kollagén – és fibrinogénkötő doméneket tartalmaz; a kollagénkötő domén (COG4932) szintén jelen van az Fmp2-ben. Javasoljuk, hogy ez a fehérje is a pilus locushoz tartozik, és nevezze el a megfelelő fmp3 gént.

Sortáz-függő pili-kódoló genomi lókuszok a finegoldia törzsek genomjában. A) az F. magna (piros) és az “F. nericia” (Világoskék) törzsek lókuszainak genomikus szervezése. A három sortáz gént Világoszöld színnel ábrázolják (“2″címkével); az fmp1 fő pilus alegység génje piros színnel (“1” jelöléssel), az fmp2 pedig narancssárga színnel (“3″jelöléssel) látható. Egyes törzsekben az fmp1-től felfelé és a sortáz génektől lefelé további gének (“8”, hemolizin III-család fehérje és “10”, hipotetikus fehérje) vannak jelen. Egy feltételezett kollagénkötő fehérje, az Fmp3 (“4” jelöléssel) az fmp1 előtt van kódolva. B) A Finegoldia törzsek Fmp1 filogenetikai fája. A legtöbb törzs egyedi Fmp1 variánssal rendelkezik. Az evolúciós történelemre a Mega7-ben végzett maximális valószínűségi módszer alkalmazásával következtettek.
az L fehérje valószínűleg az F. magna 8 legjobban vizsgált tényezője. Ez egy szuperantigén, amely a B-sejteket és az immunglobulinokat célozza meg. Az eddig vizsgált Finegoldia izolátumok körülbelül 10%-ában mutatták ki, de mérete és az antigénkötő fragmentum (Fab) kötő domének száma (B1 domén, pfam02246)4,8,18. A 312 törzsből (locus ID: M86697, 719 aa) vizsgált l fehérjével végzett robbanáskeresés néhány nagyon hasonló (50% feletti fehérje-Azonosság) fehérjét tár fel, amelyek különböző méretűek (603-1417 aa) néhány törzsben, azaz. a CCUG54800, T161024, ATCC53516, 12t273 és 12t306 törzs (ábra. 4A). Mindezek az” F. nericia ” törzsek, ami azt jelzi, hogy a B-sejtes szuperantigén gyakoribb ebben a fajban. A kivételesen nagy (1417 aa) fehérje L-szerű faktor a 12t306 törzsben két B1 domén mellett két ga modult is tartalmaz, amelyek a Peptostreptococcus albuminkötő fehérjékből ismertek (lásd alább). Számos további fehérje, amelyek alacsony hasonlósággal rendelkeznek az L fehérjével, de anélkül, hogy bármilyen látszólagos B1 domént tartalmaznának, a genomban szekvenált Finegoldia törzsekben vannak kódolva (S4A táblázat); ismét törzsspecifikus variációk léteznek, és néhány törzs, például a 09T408, a 09T494 és a T151023 nem rendelkezik ilyen homológokkal.

a Finegoldia törzsekben a protein L és FAF homológokat kódoló genomi lókuszok. (A) A 312-es törzsből (locus ID: M86697) származó, jellemzett l fehérjével végzett blasztos kereséseket genommal szekvenált Finegoldia törzsekben végeztük, és nagy hasonlóságú (>50% fehérje-Azonosság) homológokat (piros színnel, “1” jelzéssel) mutattunk be (Lásd még az S4A táblázatot). Csak” F. nericia ” törzsek, de nem F. a magna törzsek különböző hosszúságú homológokat tartalmaznak. (B) az ALB8 törzs vizsgált FAF fehérjéjével végzett blasztos keresések három F. magna (piros) és négy “F. nericia” (Világoskék) törzsben találtak teljes hosszúságú homológokat (piros, “1” jelzéssel). A FAF gént a két “F. nericia” t151023 és 12t272 törzs kereteltolja.
megállapították, hogy a Finegoldia magna kötődik az emberi albuminhoz9. Egy Peptostreptococcus albuminkötő fehérjét (PAB) vizsgáltak az ALB8 törzsben; aktivitása felgyorsult növekedési ütemhez kapcsolódik9. Az ALB8 törzs PAB fehérje (locus ID: X77864) tartalmaz GA modulokat, az albuminkötő domént; a GA modul szerkezete megoldott19. A Robbanáskeresés során kiderült, hogy a közeli PAB homológ nem található más szekvenált Finegoldia genomokban (S4B táblázat). Ehelyett több olyan homológus létezik, amelyek akár 50% – ban is megosztják a fehérje identitását; az F. magna törzsek homológjainak hasonlóságai átlagosan magasabbak, mint az “F. nericia” törzsek homológjai. Ami a ga modul tartalmú fehérjéket illeti, összesen 16 fehérjét kódolnak a 12 Finegoldia genomban, ami az albuminkötő fehérjék némi redundanciáját jelzi (S4C táblázat). A legtöbb törzs két fehérjét kódol GA modulokkal, kivéve a 09t408 (F. magna) és a T151023 (“F. nericia”) törzset.
a sejtfalhoz kötött szubtilizin-szerű proteáz SufA kimutatták, hogy lebontja számos emberi fehérjék, mint például a kollagén IV,fibrinogén és antimikrobiális peptidek,mint az LL-37 és MIG/CXCL910, 20, 21. A Blast keresések megállapították, hogy a legtöbb Finegoldia törzs SufA homológot hordoz, amely nagyon hasonló (fehérje identitás > 75%) az ALB8 törzsből származó vizsgált SufA-hoz (locus ID: DQ679960). Kivételt képeznek a 07t609, 09T408 és CCUG54800 törzsek. Egy második SufA homológ van jelen a legtöbb törzsben; átlagos fehérje identitása 26% (S4d táblázat).
egy másik fontos gazdaszervezet-interakciós tényező az FAF (F. magna adhéziós tényező). Az F. magna törzsek körülbelül 90% – a termeli ezt a faktort, amely felelős a baktériumok összetapadásáért, és a BM-4011-hez való kötődéssel közvetíti az alapmembránhoz való kötődést. Megállapítottuk, hogy a 12 törzsből hét (három F. magna és négy “F. nericia” törzs) az ALB8 törzsben tanulmányozotthoz hasonló FAF homológot kódol (ábra. 4B). Törzsspecifikus különbségek vannak, és frameshift mutációk láthatók az “F. nericia” t151023 és 12t272 törzsekben. A 09t408, 09T494 és 12t306 törzsekben nem található homológ (S4E táblázat).
a Finegoldia genomok funkcionális CAMP faktorokat kódolnak
a genomban más feltételezett gazdaszervezet-interakciós faktorokat kerestünk, és Christie-Atkins-Munch-Petersen (CAMP) faktorokat kódoló géneket találtunk. A CAMP faktorok co-hemolizinként működhetnek, és felelősek az úgynevezett CAMP reakcióért, a juh eritrociták szinergikus líziséért a Staphylococcus aureus szfingomielináz C (Béta-toxin) és a CAMP faktor22 által.
a Genombányászat két CAMP faktort kódoló gént azonosított minden genomszekvenált Finegoldia törzsben (CAMP1 és CAMP2; S4 ábra); mindegyik homológ tartalmazza a CAMP faktor család doménjét (Pfam07373). Érdekes módon törzsenként egy homológ (CAMP2) további bakteriális Ig-szerű domént (Pfam02368) tartalmaz. A CAMP1 és CAMP2 mellett négy ” F. a” nericia ” törzsek két további CAMP faktort tartalmaznak, amelyek lényegesen különböznek a többi homológtól (CAMP3 és CAMP4; S4 ábra). Így a hét “F. nericia” törzsből négy négy CAMP faktor gént tartalmaz.
ezután azt akartuk tudni, hogy a Finegoldia törzsek pozitív CAMP reakciót produkálnak-e. A CAMP agar lemez vizsgálat azt mutatja,hogy egyes törzsek erősen CAMP reakció-pozitív, míg más törzsek csak kis co-hemolízis kimutatható az alkalmazott tenyésztési körülmények között két és öt napos inkubációs (ábra. 5, S5 Ábra). Mindhárom vizsgált F. a magna törzsek erősen CAMP reakció pozitívak voltak. Között “F. nericia” törzsek kimutattunk néhány heterogenitás; két törzs erősen CAMP reakció-pozitív, négy törzs csak kismértékben co-hemolitikus, és egy törzs, CCUG54800, egyértelműen CAMP reakció-negatív. A genomikai elemzés megállapította, hogy a CCUG54800 törzsben a CAMP1-kódoló gén tartalmazott egy transzpozáz-kódoló lokusz beillesztését a gén 5′-végébe, amely megzavarja a gént (S6 ábra).

a Finegoldia törzsek CAMP reakciója. TSA agar lemezeket 5% juhvérrel használtunk. A Staphylococcus aureust a középső csíkba oltják be. A pozitív CAMP-tesztet teljes eritrocita lízis jelzi a Finegoldia sp felületén. és a S. aureus csíkok. Mindhárom F. magna (piros) törzs és a hét “F. nericia” (kék) törzs közül kettő erős pozitív CAMP reakciót mutatott két napos anaerob inkubáció után.
eredményeink azt mutatják, hogy a FINEGOLDIA SP CAMP tényezői. lehet funkcionális, és hogy vannak törzs-specifikus különbségek CAMP faktor aktivitás.
két biokémiai tesztrendszer nem képes egyértelműen megkülönböztetni az F. Magnát az “F. nericia”
GPAC biokémiai reakcióitól, beleértve a Finegoldia sp törzseket is. leírták; csak kevés biokémiai variáció a Finegoldia sp között. a törzsekről korábban beszámoltak2, 7. Két kereskedelmi forgalomban kapható biokémiai tesztrendszert teszteltünk, amelyeket az anaerobok azonosítására használnak. Az “API ons 20A” készlet 21 tesztet tartalmaz, köztük 16 tesztet a szénhidrát hasznosítására. Amint arról korábban beszámoltunk7, a finegoldia sp egyik szénhidrát sem metabolizálható. törzs (az adatok nem jelennek meg). A többi reakció (indol, ureáz, kataláz, eszkulin hidrolízis és zselatin cseppfolyósítás) szintén nem mutatott különbséget az F. magna és az “F. nericia” törzsek között (az adatok nem szerepelnek). A “RapIDTM ANA II” készlet 18 enzimatikus aktivitást tartalmaz. Míg a szacharolitikus enzimek hiányoznak, minden Finegoldia sp. a törzsek proteolitikus enzimeket termelnek (S5 táblázat). A foszfatáz aktivitást kimutató reakció volt az egyetlen reakció, amely különbözött a törzsek között, az összes F. magna törzs látszólag negatív, a legtöbb” F. nericia ” törzs pedig pozitív. Ennek a teszteredménynek az értelmezése azonban korlátozott, mivel a színváltozás a jelentőség határán volt. Összefoglalva, két létező kereskedelmi biokémiai tesztrendszer, az API 60 A és a RapIDTM ANA II, nem képes egyértelműen megkülönböztetni az F. magna-t az “F. nericia” – tól. A finegoldia SP biokémiai tulajdonságai. további biokémiai vizsgálatokkal tovább kell vizsgálni.
a mikroszkópia az F. magna és az “F. nericia”
közötti felületi különbségeket tárja fel az F. magna és az “F. nericia “pásztázó elektronmikroszkópiát (sem) és atomi erőmikroszkópiát (AFM) alkalmaztunk az F. magna (07T609, 08T492 és 09T408) és az” F. nericia ” (09T494, 12t272 és 12t306) törzseken. SEM észlelt felületi különbségek: sejtek F. magna voltak tapadó vagy aggregatív, látszólag termelő extracelluláris polimer mátrix (ábra. 6). Ez megerősíti azt a korábbi munkát, amely exopoliszacharid mátrixot vizualizált az F. magna (fmbs12 törzs) biofilmjeiben pásztázó elektronmikroszkóppal, valamint konfokális lézeres pásztázó mikroszkóppal23. A tesztelt ” F. nericia ” törzsek nem tudtunk azonosítani egy ilyen mátrixot; a sejtek kevésbé aggregatívnak tűnnek.
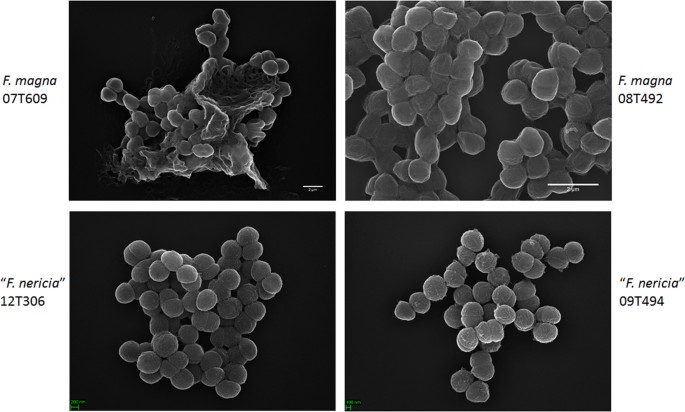
az F. magna és az “F. nericia”pásztázó elektronmikroszkópos összehasonlítása. A felső és az alsó panelek az F. magna 07t609 és 08T492, illetve az “F. nericia” 09t494 és 12t306 reprezentatív képeit mutatják. Az F. magna törzsek sejtjei jobban tapadnak / aggregálódnak, és úgy tűnik, hogy polimer mátrixot hoznak létre, különösen a 07t609 törzset; ezzel szemben az “F. nericia” sejtjei kevésbé aggregatívak.
az AFM megerősítette ezeket a megfigyeléseket. Sőt, különbségeket figyeltünk meg a sejtek függelékei tekintetében. Az F. magna sejtjei hosszabb fonalas függelékeket állítottak elő, amelyek az egész sejtfelszínről kinyúltak; ezek a struktúrák képviselhetik a pili vagy a fimbriae-t (ábra. 7). Ezzel szemben az “F. nericia” törzsek sejtjeinek simább felülete volt, hosszú kiálló függelékek nélkül. Nem zárható ki azonban, hogy ezek az AFM megfigyelések a baktériumok tenyésztési körülményeitől és a minta feldolgozási lépéseitől függenek.

atomi erő mikroszkopikus összehasonlítása F. magna és “F. nericia”. A bal és a jobb oldali panelek az F. magna 07T609 és az “F. nericia” 12t306 képeit ábrázolják. (A és C) nagyfelbontású képek támogatott fejlett soft-touch módszerrel, Z skála sávokkal akár 0,3 mm-ig és XY sávokkal, amelyek 500 nm-t jeleznek. (B és D) Overlay képek nagy kontrasztú képek (A és C), kombinálva független vízszintes és függőleges polinom eltávolítása (A és C), megoldott a 11.fokozat. A kísérleteket minden törzsből három mintán végeztük.