- Genomer Av Stammer Av Finegoldia
- Divisjon Av Finegoldia stammer i to forskjellige klader basert på genom sammenligning
- Stamme-spesifikk heterogenitet Av Finegoldia sp
- Vertsinteraksjonsfaktorer og (antatte) virulensfaktorer
- Finegoldia-genomene koder FOR CAMP-faktorer som er funksjonelle
- To biokjemiske tester systemer kan ikke entydig skille F. magna Fra «F. nericia»
- Mikroskopi avslører overflateforskjeller Mellom F. magna og «F. nericia»
Genomer Av Stammer Av Finegoldia
totalt 17 genomer Av Finegoldia sp. analyseres og sammenlignes. Ti stammer har nylig blitt sekvensert av vår gruppe15; de ble isolert fra pasienter med ortopediske leddimplantatassosierte infeksjoner i Sverige. Syv genomer ble tidligere sekvensert av andre, inkludert det lukkede genomet AV typen stamme ATCC 2932814. Genom funksjoner av alle så langt sekvenserte stammer Av Finegoldia er oppsummert I Tabell 1. Merknaden Ved Hjelp Av Prokka oppdaget mellom 1570 OG 1906 kodingssekvenser (CDER) per genom, med et gjennomsnitt på 1760 CDER per stamme.
Divisjon Av Finegoldia stammer i to forskjellige klader basert på genom sammenligning
vi har tidligere lagt merke til en stor heterogenitet blant genomer Av Finegoldia isolater15. En kjerne-genom justering av alle 17 genomer ble gjort og enkelt nukleotid polymorfismer (SNPs) i kjernen genomet ble kalt. Totalt ble 126 647 kjerne-genom Snper identifisert Og brukt til fylogenomisk rekonstruksjon, og avslørte at de 17 genomene kan grupperes i to forskjellige klader (Fig. 1). En ytterligere analyse som bestemmer gjennomsnittlig nukleotididentifikasjon (ANI) av kjernegenomet viste at de to kladene utviser EN ANI på 90,7%, med en stamme-spesifikk ANI-variasjon fra 90,2% til 91,2%. En slik lav ANI indikerer at de to clades representerer individuelle arter16. En klade inneholder de fleste tidligere sekvenserte og studerte stammer, inkludert ATCC29328 OG ALB8; det kan betraktes som den klassiske F. magna clade, med en intraklade ANI på 96,6% (variasjon 95,1-97,4%). Den andre kladen med en intraklade ANI på 94,8% (93,4-96.6%) inneholder en annen ATCC-stamme (ATCC 53516) og hovedsakelig stammer isolert fra pasienter ved ③rebro Universitetssykehus, Sweden15. Heretter er stammer av denne kladen foreløpig kalt «Finegoldia nericia» (Latin For nä, den svenske regionen, hvor de fleste stammer av denne arten ble isolert).

Fylogenomisk tre basert på kjerne-genom SNPs av alle 17 sekvenserte genomer Av Finegoldia stammer. Programmet Parsnp ble brukt til å justere kjernegenomet og ringe SNPs. Kjernegenomet er representert av 43% av referansegenomet (ATCC29328); totalt 126 647 pålitelige kjerne-genom Snper ble brukt til å rekonstruere et helgenomfylogeni. Genomer kan tildeles to hoved clades; en clade representerer F. magna stammer og den andre clade består av stammer av en ny art, her forsøksvis kalt «F. nericia».
en sammenligning AV 16s rRNA-sekvenser av alle isolater viste en høy grad av total likhet (>99%), men oppdaget en separasjon Av F. magna-isolater fra» F. nericia «- isolater, på grunn av 5 til 9 » F. nericia » – spesifikke SNPs (Figur S1). Det viste også en høyere diversifisering blant» F. nericia » isolater sammenlignet Med F. magna. Deretter analyserte vi ytterligere 16s rRNA-sekvenser som tilhører Finegoldia sp. lagret I GenBank (Figur S2). Dette viste at de fleste lagrede sekvenser klynger seg med «F. nericia», spesielt En gruppe Finegoldia sp. stammer isolert fra biliær stent biofilmer17. Igjen oppdages en høyere diversifisering blant» F. nericia » isolater. Deretter analyserte vi humane mikrobiomdata lagret I img (Integrated Microbial Genomes and microbiome samples) database for tilstedeværelse Av f. magna og «F. nericia» sekvenser. 3287 og 7068 kodende sekvenser Av Henholdsvis F. magna ATCC29328 og «F. nericia» atcc 53516 kunne identifiseres i de 875 samlede humane mikrobiomdatasettene som er tilstede I IMG( data ikke vist); høyeste forekomster Av Finegoldia ble påvist i retroaurikulære krøllprøver. Samlet sett tyder disse analysene på at blant menneskelige assosierte Finegoldia isolater «F. nericia» synes å være mer utbredt Enn F. magna.
Stamme-spesifikk heterogenitet Av Finegoldia sp
for å kaste ytterligere lys på befolkningsstrukturen I finegoldia slekten, komparative analyser av 12 genomer (alle ti nylig sekvenserte stammer og de to atcc stammer) ble utført, inkludert fire stammer Av F. magna clade og åtte stammer av «F. nericia» clade. En visuell sammenligning av de 12 genomene fremhevet nukleotididentitetsforskjellen mellom de to kladene(Fig. 2). Interessant synes plasmidet funnet i ATCC 29328 å være spesifikt for denne stammen, da ingen annen sekvensert stamme inneholder en lignende sekvens(Fig . 2A).

Genom sammenligning av 12 stammer som tilhører slekten Finegoldia. De to innerste ringene representerer G + C-innholdet (svart) og GC-skjevheten (fiolett/grønn). (A) referansestammen er ATCC29328( F. magna); merk den høye nukleotididentiteten (visualisert av den mørke røde fargen) av de tre innerste genomene (som tilhører Alle Til F. «F. nericia «(blek rød farge på de åtte ytre ringene); fra innsiden til utsiden: stammer 07T609, 08t492, 09t408, 09T494, 12t272, 12t273, 12T306, CCUG 54800, T151023, T160124 OG ATCC 53516. Den store atcc29328-spesifikke regionen i øvre venstre region representerer plasmid pFMC. (B) referansestammen ER 09T494 («F. nericia»). De syv» F. nericia » – genomene (indre ringer) har en høyere nukleotididentifikasjon (representert ved mørkere rød farge), sammenlignet med de fire F. magna-genomene( ytre ringer); fra innsiden til utsiden: stammer 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 OG 09T408. Det er ingen tegn til «F. nericia» – spesifikke genomiske regioner, men merk de fem store stamme – spesifikke klynger.
ingen signifikante forskjeller i genomstørrelse og antall CDER ble oppdaget mellom de to clades. I tillegg ble ingen clade-spesifikke (dvs. f. magna eller «F. nericia»-spesifikke) genomiske regioner detektert, som dømt fra BRIG-analysen (Fig. 2) og også bekreftet av en toveis blast tilnærming av ALLE CDER bruker ProteinOrtho (Tabell S1). Denne tilnærmingen oppdaget 1202 orthologer delt av alle 12 genomer, dvs. i gjennomsnitt er 68% AV CDENE av hver stamme en del av kjerneproteomet. Bare svært få clade-spesifikke gener er funnet. I stedet kan mange regioner deles av bare noen få stammer, f. eks for stamme 09T494 (Se Også Fig. 2B). I tillegg er stamme-spesifikke regioner til stede som koder i totalt 1016 stamme-spesifikke CDER, med et stort område mellom stammene, f. eks 187 og 22 stamme-spesifikke CDER er påvist i stamme ATCC29328 og t151023, henholdsvis (Tabell S1). Blant de belastningsspesifikke funksjonene er ulike transportfunksjoner (jern, efflukspumper, oligopeptid ABC-transportører), overflateproteiner, restriksjonsmodifiseringssystemer, konjugative overføringsfunksjoner, polyketidsyntese, bakteriociner, fagrelaterte funksjoner, antibiotikaresistensdeterminanter og CRISPR/cas-systemer.
Interessant ble forskjeller I CRISPR / cas-systemene detektert mellom stammer Av F. magna og «F. nericia» (Tabell S2). Syv Av ni F. magna stammer inneholdt en til fire CRISPR arrays per stamme, som inneholdt i gjennomsnitt 12 avstandsstykker (fra 3 til 38 avstandsstykker per stamme). Gjenta «GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC» var spesifikk for stammer Av F. magna. I motsetning til dette hadde «F. nericia» – stammer bare en CRISPR-array per stamme som inneholdt gjennomsnittlig 53 avstandsstykker (område 14 til 87 per stamme). Hver stamme inneholdt et individuelt sett av avstandsstykker i DERES CRISPR array( er), understreker stamme-spesifikk heterogenitet, og indikerer at hver stamme har en unik evolusjonær historie.
for å vurdere om slike belastningsspesifikke funksjoner ble horisontalt anskaffet, så vi på tegn på mobilitet / oppkjøp ved Hjelp Av IslandViewer-programmet (Figur S3). Dette programmet forutslo 141 (stamme 09T408) til 408 (stamme 12T273) gener som ligger i genomiske øyer som sannsynligvis vil bli horisontalt anskaffet, inkludert mange av de ovennevnte stammespesifikke gener involvert i spesialisering, fitness, overlevelse og interspecies konkurranse (Tabell S3).
Vertsinteraksjonsfaktorer og (antatte) virulensfaktorer
Flere vertsinteraksjonsfaktorer er beskrevet For F. magna 4,5. Sortase-avhengig pili har blitt identifisert som er viktig for adhesjon og kolonisering13. Et genomisk locus som koder for gener for pilus-underenheten Fmp1, den antatte underenheten Fmp2 og tre sortaser er bevart blant alle sekvenserte genomer (Fig . 3A). Det er belastningsspesifikke variasjoner opp-og nedstrøms for dette lokus, dvs. noen stammer har flere gener oppstrøms for fmp1 og nedstrøms for sortase locus, inkludert et gen som koder for et hemolysin III familieprotein. Interessant er det belastningsspesifikke forskjeller angående fmp1. Nesten hver stamme bærer en individuell variant og en fylogenetisk analyse Av Fmp1 på proteinnivå viste ingen korrelasjon med fylogenomisk analyse av kjernegenomet (Fig. 3B). I motsetning Er Fmp2 svært bevart blant alle stammer. Et annet protein, kodet oppstrøms av fmp1, har kollagen – og fibrinogenbindende domener; det kollagenbindende domenet (COG4932) er også til stede I Fmp2. Vi foreslår at dette proteinet også tilhører pilus locus og heter det tilsvarende genet fmp3.

Sortase-avhengig pili-koding genomisk loci i genomene Av Finegoldia stammer. (A) Genomisk organisering av loci I F. magna (rød) og «F. nericia» (lyseblå) stammer. De tre sortasegenene er avbildet i lysegrønn (merket «2»); genet For den store pilus-underenheten Fmp1 er vist i rødt (merket «1») og fmp2 er avbildet i oransje (merket «3»). Oppstrøms for fmp1 og nedstrøms for sortasegenene i noen stammer finnes ytterligere gener (merket «8», hemolysin III-familieprotein og «10», hypotetisk protein). Et antatt kollagenbindende protein, Fmp3 (merket «4»), er kodet oppstrøms fmp1. (B) Fylogenetisk tre Av Fmp1 Av Finegoldia stammer. De fleste stammer har en individuell fmp1-variant. Den evolusjonære historien ble utledet ved Å bruke Maksimal Sannsynlighetsmetode, gjort I MEGA7.
Protein L er trolig Den best studerte faktoren Av f. magna 8. Det er et superantigen som er rettet Mot B-celler og immunglobuliner. Det har blitt påvist i ca 10% Av De hittil testede finegoldia isolater, men varierer i størrelse, og antall antigen-bindende fragment (Fab) – bindende domener (B1 domene, pfam02246)4,8,18. Et blastsøk med det studerte proteinet L fra stamme 312 (locus ID: M86697, 719 aa) avslører noen få svært like (proteinidentitet over 50%) proteiner med varierende størrelser (603 til 1417 aa) i noen få stammer, dvs. stammer CCUG54800, T161024, ATCC53516, 12T273 OG 12t306 (Fig. 4A). Alle disse er» F. nericia » – stammer, noe som indikerer At b-celle superantigen er mer utbredt i denne arten. Den eksepsjonelt store (1417 aa) protein L-lignende faktor i stamme 12T306 besitter foruten to b1 domener også TO GA moduler som er kjent fra peptostreptokokk albumin-bindende proteiner (se nedenfor). Mange ekstra proteiner med lav likhet med protein L, men uten å inneholde noen tilsynelatende b1 domener er kodet I Genomet-sekvenserte Finegoldia stammer (Tabell S4a); igjen eksisterer stamme – spesifikke variasjoner, og NOEN få stammer, dvs. 09T408, 09T494 Og T151023 har ingen slike homologer.

Genomisk loci som koder for protein l og FAF-homologer I Finegoldia-stammer. (A) Blastsøk med det karakteriserte proteinet L fra stamme 312 (locus ID: M86697) ble utført I genomsekvenserte Finegoldia-stammer, og homologer (i rødt, merket «1») med høy likhet (>50% proteinidentitet) er vist (Se Også Tabell S4A). Bare «F. nericia» stammer, Men Ikke F. magna stammer inneholder slike homologer med varierende lengder. (B) Blastsøk med DET studerte FAF-proteinet AV stamme ALB8 fant full lengde homologer (i rødt, merket «1») i tre F. magna (rød) og fire «F. nericia» (lyseblå) stammer. FAF-genet er frameshifted i de to» F. nericia » – stammene T151023 Og 12t272.
Finegoldia magna ble funnet å binde seg til humant albumin 9. Et peptostreptokokkalbuminbindende protein (PAB)er studert i stammen ALB8; dets aktivitet er knyttet til en akselerert vekstrate9. PAB-proteinet AV stamme ALB8 (locus ID: X77864) inneholder ga-moduler, albumin-bindende domene; strukturen TIL ga-modulen har blitt løst19. Et Blastsøk viste at en nær pab homolog ikke kan bli funnet i andre sekvenserte Finegoldia-genomer (Tabell S4B). I stedet er det flere homologer som deler opptil 50% proteinidentitet; likheter av homologer I F. magna-stammer er i gjennomsnitt høyere sammenlignet med homologer i» F. nericia » – stammer. Når DET gjelder GA-modulholdige proteiner, er totalt 16 proteiner kodet i 12 Finegoldia-genomene, noe som indikerer noe redundans av albumin-bindende proteiner (Tabell S4C). De fleste stammer koder to proteiner MED GA-moduler, bortsett FRA stamme 09T408 (F. magna) Og T151023 («F. nericia»).
det celleveggfestede subtilisinlignende protease SufA har vist seg å nedbryte flere humane proteiner, som kollagen IV, fibrinogen og antimikrobielle peptider SOM LL-37 og MIG/CXCL910,20,21. Blast søk fastslått at De Fleste Finegoldia stammer bære En SufA homolog som er svært lik (protein identitet > 75%) til den studerte SufA fra stamme ALB8 (locus ID: DQ679960). Unntak er stammene 07T609, 09T408 OG CCUG54800. En annen sufa homolog er tilstede i de fleste stammer; den har en gjennomsnittlig proteinidentitet på 26% (Tabell S4D).
EN annen viktig vertsinteraksjonsfaktor ER FAF (F. magna adhesjonsfaktor). Rundt 90% Av F. magna-stammer produserer denne faktoren, som er ansvarlig for klumping av bakterier og medierer binding til kjellermembranen ved binding TIL BM-4011. Vi fant at syv (tre F. magna og fire «F. nericia» stammer) ut av de 12 stammene koder FOR EN FAF homolog som ligner den som ble studert I ALB8-stammen (Fig. 4B). Stamme – spesifikke forskjeller eksisterer og rammeskift mutasjoner er sett i» F. nericia » stammer T151023 OG 12t272. Ingen homolog finnes I stammene 09T408, 09T494 OG 12T306 (Tabell S4E).
Finegoldia-genomene koder FOR CAMP-faktorer som er funksjonelle
vi søkte i genomet etter andre antatte vertsinteraksjonsfaktorer og fant gener som koder For Christie-Atkins-Munch-Petersen (CAMP) – faktorer. CAMP-faktorer kan fungere som co-hemolysiner og står for den såkalte CAMP-reaksjonen, den synergistiske lysis av sau erythrocytter Av Staphylococcus aureus sphingomyelinase C (beta-toksin) og EN CAMP-faktor22.
Genomutvinning identifiserte to gener som koder FOR CAMP-faktorer i hver Genom-sekvensert Finegoldia-stamme (CAMP1 OG CAMP2; Figur S4); hver homolog har CAMPFAKTORFAMILIEDOMENET (Pfam07373). Interessant, en homolog (CAMP2) per stamme havner et ekstra bakterielt Ig – lignende domene (Pfam02368). I TILLEGG TIL CAMP1 OG CAMP2, fire » F. nericia » stammer inneholder to EKSTRA CAMP faktorer som vesentlig skiller seg fra de andre homologs (CAMP3 OG CAMP4; Figur S4). Dermed har fire av syv «F. nericia» – stammer fire CAMPFAKTORGENER.
deretter ønsket Vi å vite Om Finegoldia-stammer gir en positiv CAMPREAKSJON. EN CAMP agar plate analyse viser at noen stammer er sterkt CAMP reaksjon-positive, mens for andre stammer bare lite co-hemolyse kan påvises under de anvendte dyrkningsbetingelser på to og fem dager med inkubasjon (Fig. 5, Figur S5). Alle tre testet F. magna stammer var sterkt CAMP reaksjon-positive. Blant» F. nericia » – stammer oppdaget vi noe heterogenitet; to stammer var sterkt CAMP-reaksjon-positive, fire stammer var bare litt co-hemolytiske, og EN stamme, CCUG54800, var tydelig CAMP-reaksjon-negativ. Genomisk analyse fant at i stamme CCUG54800 INNEHOLDT CAMP1-kodende genet en innføring av et transposase-kodende locus i 5 ‘ – enden av genet, som forstyrrer genet (Figur S6).

CAMP reaksjon Av Finegoldia stammer. TSA agar plater med 5% sau blod ble brukt. Staphylococcus aureus er inokulert i midtstripen. En positiv CAMP-test er indikert ved fullstendig erytrocytlysis ved grensesnittet Til Finegoldia sp. Og S. aureus-stripene. Alle Tre F. magna (i rødt) stammer og to av syv «F. nericia» (i blått) stammer viste en sterk positiv CAMP reaksjon etter to dager med anaerob inkubasjon.
Våre resultater indikerer AT CAMP faktorer Av Finegoldia sp. kan være funksjonell og at det er belastningsspesifikke forskjeller I CAMPFAKTORAKTIVITET.
To biokjemiske tester systemer kan ikke entydig skille F. magna Fra «F. nericia»
Biokjemiske reaksjoner AV GPAC inkludert stammer Av Finegoldia sp. har blitt beskrevet; bare lite biokjemisk variasjon Blant Finegoldia sp. stammer ble tidligere rapportert2, 7. Vi testet to kommersielt tilgjengelige biokjemiske testsystemer som brukes til identifisering av anaerober. Settet «API® 20A» inneholder 21 tester, blant dem 16 tester for karbohydratutnyttelse. Som rapportert tidligere7, kunne Ingen av karbohydrater metaboliseres Av Noen Finegoldia sp. stamme(data ikke vist). De andre reaksjonene (indol, urease, katalase, esculinhydrolyse og gelatinfortynning) viste heller ingen forskjell mellom stammer Av F. magna og «F. nericia» (data ikke vist). Settet «RapIDTM ANA II» inneholder 18 tester for enzymatiske aktiviteter. Mens sakkarolytiske enzymer mangler, Alle Finegoldia sp. stammer produserer proteolytiske enzymer (Tabell S5). Reaksjonen som detekterte fosfataseaktivitet var den eneste reaksjonen som var forskjellig blant stammene, med Alle F. magna-stammer tilsynelatende negative og de fleste» F. nericia » – stammer positive. Imidlertid er tolkningen av dette testresultatet begrenset siden fargeendringen var på grensen av betydning. Til slutt kan to eksisterende kommersielle biokjemiske testsystemer, API® 20 A og RapIDTM ANA II, ikke entydig skille F. magna fra»F. nericia». De biokjemiske egenskapene Til Finegoldia sp. trenger å bli ytterligere utforsket med flere biokjemiske tester.
Mikroskopi avslører overflateforskjeller Mellom F. magna og «F. nericia»
i et forsøk på å visualisere morfologi og overflatestruktur Av f. magna og » F. nericia «vi brukte scanning elektronmikroskopi (SEM) og atomic force mikroskopi (AFM) på stammer Av F. magna (07T609, 08T492, OG 09T408) og» F. nericia » (09T494, 12t272 OG 12T306). Sem oppdaget overflateforskjeller: celler Av F. magna var mer adherente eller aggregative, tilsynelatende produserer en ekstracellulær polymermatrise (Fig. 6). Dette bekrefter tidligere arbeid som har visualisert en eksopolysakkaridmatrise i biofilmer Av F. magna (stamme FmBs12) ved å skanne elektronmikroskopi samt konfokal laserskanning mikroskopi23. I den testede » F. nericia » stammer vi ikke kunne identifisere en slik matrise; celler virker mindre aggregative.
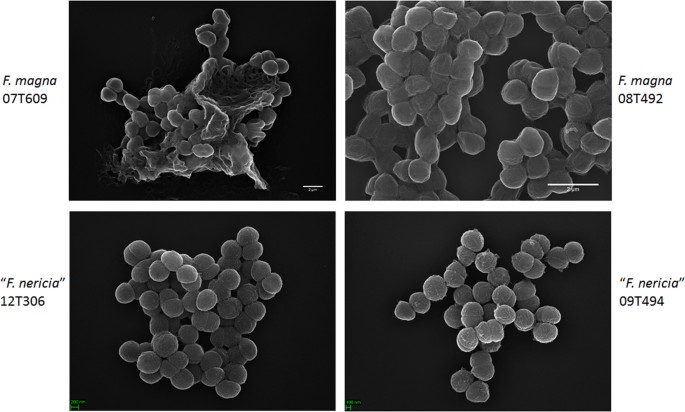
Scanning elektronmikroskopisk sammenligning Av F. magna og «F. nericia». De øvre og nedre panelene viser representative bilder av Henholdsvis F. magna 07t609 og 08t492 og «F. nericia» 09T494 og 12t306. Celler Av F. magna-stammer er mer adherente/aggregative og synes å produsere en polymermatrise, spesielt stamme 07T609; i kontrast er celler av «F. nericia» mindre aggregative.
AFM bekreftet disse observasjonene. Videre observerte vi forskjeller angående cellebidrag. Celler Av F. magna produserte lengre filamentøse appendager som stakk ut fra hele celleoverflaten; disse strukturene kunne representere pili eller fimbriae (Fig. 7). I kontrast hadde celler av «F. nericia» – stammer en jevnere overflate uten lange fremspringende vedlegg. Det kan imidlertid ikke utelukkes at DISSE AFM-observasjonene avhenger av bakteriekultiveringsforholdene og prøvebehandlingstrinnene.

Atomic force mikroskopisk sammenligning Av F. magna og «F. nericia». Venstre og høyre paneler representerer bilder av Henholdsvis F. magna 07T609 og «F. nericia» 12t306. (A og C) høyoppløselige bilder med assistert avansert soft-touch metode Med Z skala barer opp til 0,3 µ og XY barer som indikerer 500 nm. (B og D) Overlay bilder av høykontrastbilder av (A Og C), kombinert med uavhengig horisontal og vertikal polynomisk fjerning av (A Og C), løst til 11. grad. Eksperimenter ble utført på tre prøver fra hver stamme.