- Genome von Stämmen von Finegoldia
- Aufteilung der Finegoldia-Stämme in zwei verschiedene Kladen basierend auf dem Genomvergleich
- Stammspezifische Heterogenität von Finegoldia sp
- Wirt-wechselwirkende Faktoren und (mutmaßliche) Virulenzfaktoren
- Finegoldia-Genome kodieren CAMP-Faktoren, die funktionell sind
- Zwei biochemische Testsysteme können F. magna nicht eindeutig von „F. nericia“ unterscheiden
- Mikroskopie zeigt Oberflächenunterschiede zwischen F. magna und „F. nericia“
Genome von Stämmen von Finegoldia
Insgesamt 17 Genome von Finegoldia sp. wurden analysiert und verglichen. Zehn Stämme wurden kürzlich von unserer Gruppe sequenziert15; Sie wurden von Patienten mit orthopädischen Gelenkimplantat-assoziierten Infektionen in Schweden isoliert. Sieben Genome wurden zuvor von anderen sequenziert, darunter das geschlossene Genom des Typstamms ATCC 2932814. Genommerkmale aller bisher sequenzierten Stämme von Finegoldia sind in Tabelle 1 zusammengefasst. Die Annotation mit Prokka detektiert zwischen 1570 und 1906 kodierende Sequenzen (CDS) pro Genom, mit einem Durchschnitt von 1760 CDS pro Stamm.
Aufteilung der Finegoldia-Stämme in zwei verschiedene Kladen basierend auf dem Genomvergleich
Wir haben zuvor eine große Heterogenität unter den Genomen von Finegoldia-Isolaten festgestellt15. Eine Kerngenom-Ausrichtung aller 17 Genome wurde durchgeführt und Einzelnukleotidpolymorphismen (SNPs) im Kerngenom wurden aufgerufen. Insgesamt wurden 126.647 Kerngenom-SNPs identifiziert und für die phylogenomische Rekonstruktion verwendet, was zeigt, dass die 17 Genome in zwei verschiedene Kladen gruppiert werden können (Abb. 1). Eine zusätzliche Analyse, die die durchschnittliche Nukleotiddichte (ANI) des Kerngenoms bestimmt, zeigte, dass die beiden Kladen eine ANI von 90,7% aufweisen, mit einer stammspezifischen ANI-Variation von 90,2% bis 91,2%. Ein derart niedriger ANI zeigt an, dass die beiden Kladen einzelne Arten repräsentieren16. Eine Klade enthält die meisten zuvor sequenzierten und untersuchten Stämme, einschließlich ATCC29328 und ALB8; Es kann als die klassische F. magna-Klade angesehen werden, mit einem Intrakladenani von 96,6% (Variation 95,1–97,4%). Die andere Klade mit einem Intrakladenani von 94,8% (93,4-96.6%) enthält einen weiteren ATCC-Stamm (ATCC 53516) und hauptsächlich Stämme, die von Patienten des Universitätsklinikums Örebro, Schweden, isoliert wurden15. Im Folgenden werden Stämme dieser Klasse vorläufig „Finegoldia nericia“ genannt (lateinisch für Närke, die schwedische Region, in der die meisten Stämme dieser Art isoliert wurden).

Phylogenomischer Baum basierend auf Kerngenom-SNPs aller 17 sequenzierten Genome von Finegoldia-Stämmen. Das Programm Parsnp wurde verwendet, um das Kerngenom auszurichten und SNPs aufzurufen. Das Kerngenom wird durch 43% des Referenzgenoms (ATCC29328) repräsentiert; Insgesamt wurden 126.647 zuverlässige Kerngenom-SNPs verwendet, um eine Phylogenie des gesamten Genoms zu rekonstruieren. Genome können zwei Hauptklassen zugeordnet werden; Eine Klasse repräsentiert F. magna-Stämme und die andere Klasse besteht aus Stämmen einer neuen Art, hier vorläufig „F. nericia“ genannt.
Ein Vergleich von 16S-rRNA-Sequenzen aller Isolate ergab einen hohen Grad an Gesamtähnlichkeit (> 99%), zeigte jedoch eine Trennung von F. magna-Isolaten von „F. nericia“ -Isolaten aufgrund von 5 bis 9 „F. nericia“-spezifische SNPs (Abbildung S1). Es zeigte sich auch eine höhere Diversifikation unter „F. nericia“ -Isolaten im Vergleich zu F. magna. Als nächstes analysierten wir zusätzliche 16S-rRNA-Sequenzen von Finegoldia sp. gespeichert in GenBank (Abbildung S2). Dies zeigte, dass die meisten gespeicherten Sequenzen Cluster mit „F. nericia“, insbesondere eine Gruppe von Finegoldia sp. stämme, die aus biliären Stent-Biofilmen isoliert wurden17. Auch hier wird eine höhere Diversifikation unter „F. nericia“ -Isolaten nachgewiesen. Als nächstes analysierten wir humane Mikrobiomdaten, die in der IMG-Datenbank (Integrated Microbial Genomes and microbiome samples) gespeichert sind, auf das Vorhandensein von F. magna- und „F. nericia“ -Sequenzen. 3287 bzw. 7068 kodierende Sequenzen von F. magna ATCC29328 bzw. „F. nericia“ ATCC 53516 konnten in den 875 im IMG vorliegenden humanen Mikrobiom-Datensätzen identifiziert werden (Daten nicht dargestellt); höchste Vorkommen von Finegoldia wurden in retroaurikulären Faltenproben nachgewiesen. Zusammengenommen deuten diese Analysen darauf hin, dass „F. nericia“ unter den humanassoziierten Finegoldia-Isolaten häufiger zu sein scheint als F. magna.
Stammspezifische Heterogenität von Finegoldia sp
Um die Populationsstruktur der Gattung Finegoldia weiter zu beleuchten, wurden vergleichende Analysen von 12 Genomen (alle zehn neu sequenzierten Stämme und die beiden ATCC-Stämme) durchgeführt, darunter vier Stämme der F. magna-Clade und acht Stämme der „F. nericia“-clade. Ein visueller Vergleich der 12 Genome hob den Nucleotid-Identitätsunterschied zwischen den beiden Kladen hervor (Abb. 2). Interessanterweise scheint das in ATCC 29328 gefundene Plasmid für diesen Stamm spezifisch zu sein, da kein anderer sequenzierter Stamm eine ähnliche Sequenz enthält (Abb. 2A).

Genomvergleich von 12 Stämmen der Gattung Finegoldia. Die beiden innersten Ringe repräsentieren den G+C-Gehalt (schwarz) und den GC-Skew (violett/grün). (A) Der Referenzstamm ist ATCC29328 (F. magna); Beachten Sie die hohe Nukleotididentität (sichtbar durch die dunkelrote Farbe) der drei innersten Genome (die alle zu F gehören. magna), und die unteren identifizieren sich mit acht Stämmen, die zu „F. nericia“ gehören (blassrote Farbe der acht äußeren Ringe); von innen nach außen: Stämme 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 und ATCC 53516. Die große ATCC29328-spezifische Region in der oberen linken Region repräsentiert das Plasmid pFMC. B) Der Referenzstamm ist 09T494 („F. nericia“). Die sieben „F. nericia“ -Genome (Innenringe) haben eine höhere Nukleotidkonzentration (dargestellt durch die dunklere rote Farbe) als die vier F. magna-Genome (Außenringe); von innen nach außen: stämme 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 und 09T408. Es gibt keine Anzeichen für „F. nericia“ -spezifische genomische Regionen, aber beachten Sie die fünf großen stammspezifischen Cluster.
Es wurden keine signifikanten Unterschiede in der Genomgröße und Anzahl der CDS zwischen den beiden Kladen festgestellt. Darüber hinaus wurden keine kladenspezifischen (d. h. F. magna- oder „F. nericia“ -spezifischen) genomischen Regionen nachgewiesen, wie aus der BRIG-Analyse (Abb. 2) und auch durch einen bidirektionalen Blastansatz aller CDS mit ProteinOrtho bestätigt (Tabelle S1). Bei diesem Ansatz wurden 1202 Orthologe nachgewiesen, die von allen 12 Genomen gemeinsam genutzt werden, d. h. Im Durchschnitt sind 68% der CDS jedes Stammes Teil des Kernproteoms. Es werden nur sehr wenige kladenspezifische Gene gefunden. Stattdessen finden sich viele Regionen, die nur wenigen Stämmen gemeinsam sind, z.B. für Stamm 09T494 (siehe auch Abb. 2B). Zusätzlich sind stammspezifische Bereiche vorhanden, die insgesamt 1016 stammspezifische CDS kodieren, mit einem großen Bereich zwischen den Stämmen, z.B. 187 und 22 stammspezifische CDS werden in Stamm ATCC29328 bzw. T151023 detektiert (Tabelle S1). Zu den stammspezifischen Funktionen gehören verschiedene Transportfunktionen (Eisen, Effluxpumpen, Oligopeptid-ABC-Transporter), Oberflächenproteine, Restriktionsmodifikationssysteme, konjugative Transferfunktionen, Polyketidsynthese, Bakteriocine, phagenbezogene Funktionen, Antibiotikaresistenzdeterminanten und CRISPR /cas-Systeme.
Interessanterweise wurden Unterschiede in den CRISPR/cas-Systemen zwischen Stämmen von F. magna und „F. nericia“ nachgewiesen (Tabelle S2). Sieben von neun F. magna-Stämme enthielten ein bis vier CRISPR-Arrays pro Stamm, die durchschnittlich 12 Abstandshalter enthielten (von 3 bis 38 Abstandshalter pro Stamm). Die Wiederholung „GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC“ war spezifisch für Stämme von F. magna. Im Gegensatz dazu trugen „F. nericia“ -Stämme nur ein CRISPR-Array pro Stamm, das durchschnittlich 53 Spacer enthielt (Bereich 14 bis 87 pro Stamm). Jeder Stamm enthielt einen individuellen Satz von Spacern in seinen CRISPR-Arrays, was die stammspezifische Heterogenität untermauerte und darauf hinwies, dass jeder Stamm eine einzigartige Evolutionsgeschichte hat.
Um zu beurteilen, ob solche dehnungsspezifischen Funktionen horizontal erfasst wurden, untersuchten wir die Anzeichen ihrer Mobilität / Erfassung mit dem IslandViewer-Programm (Abbildung S3). Dieses Programm prognostizierte 141 (Stamm 09T408) bis 408 (Stamm 12T273) Gene auf genomischen Inseln, die wahrscheinlich horizontal erworben werden, einschließlich vieler der oben genannten stammspezifischen Gene, die an Spezialisierung, Fitness, Überleben und Interspezies-Wettbewerb beteiligt sind (Tabelle S3).
Wirt-wechselwirkende Faktoren und (mutmaßliche) Virulenzfaktoren
Für F. magna 4,5 wurden mehrere Wirt-wechselwirkende Faktoren beschrieben. Es wurden Sortase-abhängige Pili identifiziert, die für die Adhäsion und Besiedlung wichtig sind13. Ein genomischer Locus, der die Gene für die Pilus-Untereinheit Fmp1, die mutmaßliche Untereinheit Fmp2 und drei Sortasen kodiert, ist unter allen sequenzierten Genomen konserviert (Abb. 3A). Es gibt dehnungsspezifische Variationen up- und downstream dieses Locus, d.h. einige Stämme besitzen zusätzliche Gene stromaufwärts von fmp1 und stromabwärts des Sortase-Locus, einschließlich eines Gens, das für ein Protein der Hämolysin-III-Familie kodiert. Interessanterweise gibt es stammspezifische Unterschiede in Bezug auf fmp1. Fast jeder Stamm trägt eine individuelle Variante und eine phylogenetische Analyse von Fmp1 auf Proteinebene zeigte keine Korrelation mit der phylogenomischen Analyse des Kerngenoms (Abb. 3B). Im Gegensatz dazu ist Fmp2 unter allen Stämmen hoch konserviert. Ein anderes Protein, das stromaufwärts von fmp1 kodiert ist, beherbergt Kollagen- und Fibrinogenbindungsdomänen; die Kollagen-bindende Domäne (COG4932) ist auch in Fmp2 vorhanden. Wir schlagen vor, dass dieses Protein auch zum Pilus-Locus gehört und nennen das entsprechende Gen fmp3.

Sortase-abhängige Pili-kodierende genomische Loci in den Genomen von Finegoldia-Stämmen. (A) Genomische Organisation der Loci in F. magna (rot) und „F. nericia“ (hellblau) Stämmen. Die drei Sortasen-Gene sind hellgrün dargestellt (beschriftet mit „2“); das Gen für die große Pilus-Untereinheit Fmp1 ist rot (mit „1“ gekennzeichnet) und fmp2 orange (mit „3“ gekennzeichnet) dargestellt. Stromaufwärts von fmp1 und stromabwärts der Sortase-Gene sind in einigen Stämmen zusätzliche Gene (markiert „8“, Hämolysin III-Familienprotein und „10“, hypothetisches Protein) vorhanden. Ein mutmaßliches Kollagen-bindendes Protein, Fmp3 (markiert „4“), ist stromaufwärts von fmp1 codiert. (B) Phylogenetischer Baum von Fmp1 von Finegoldia-Stämmen. Die meisten Stämme besitzen eine individuelle Fmp1-Variante. Die Evolutionsgeschichte wurde mit der Maximum-Likelihood-Methode abgeleitet, die in MEGA7 durchgeführt wurde.
Protein L ist wahrscheinlich der am besten untersuchte Faktor von F. magna 8. Es ist ein Superantigen, das auf B-Zellen und Immunglobuline abzielt. Es wurde in etwa 10% der bisher getesteten Finegoldia-Isolate nachgewiesen, variiert jedoch in Größe und Anzahl der Antigen-bindenden Fragment (Fab)-bindenden Domänen (B1-Domäne, pfam02246)4,8,18. Eine Blastsuche mit dem untersuchten Protein L aus Stamm 312 (Locus ID: M86697, 719 aa) zeigt in einigen Stämmen einige sehr ähnliche (Proteinidentität über 50%) Proteine mit unterschiedlichen Größen (603 bis 1417 aa), d.h. stämme CCUG54800, T161024, ATCC53516, 12T273 und 12T306 (Abb. 4A). All dies sind „F. nericia“ -Stämme, was darauf hindeutet, dass das B-Zell-Superantigen bei dieser Spezies häufiger vorkommt. Der außergewöhnlich große (1417 aa) Protein-L-like-Faktor im Stamm 12T306 besitzt neben zwei B1-Domänen auch zwei GA-Module, die von Peptostreptokokken-Albumin-bindenden Proteinen bekannt sind (siehe unten). In den genomsequenzierten Finegoldia-Stämmen sind viele zusätzliche Proteine mit geringer Ähnlichkeit zu Protein L codiert, die jedoch keine offensichtlichen B1-Domänen enthalten (Tabelle S4A); auch hier existieren stammspezifische Variationen, und einige Stämme, d.h. 09T408, 09T494 und T151023 besitzen keine solchen Homologen.

Genomische Loci, die Protein L- und FAF-Homologe in Finegoldia-Stämmen kodieren. (A) In genomsequenzierten Finegoldia-Stämmen wurden Blastenrecherchen mit dem charakterisierten Protein L aus Stamm 312 (Locus ID: M86697) durchgeführt und Homologe (in rot, mit „1“ gekennzeichnet) mit hoher Ähnlichkeit (>50% Proteinidentität) gezeigt (siehe auch Tabelle S4A). Nur „F. nericia“ -Stämme, aber nicht F. magna-Stämme enthalten solche Homologen mit unterschiedlichen Längen. (B) Blastrecherchen mit dem untersuchten FAF-Protein des Stammes ALB8 fanden in drei F. magna (rot) und vier „F. nericia“ (hellblau) Stämmen Homologe in voller Länge (in rot, mit „1“ gekennzeichnet). Das FAF-Gen ist in den beiden „F. nericia“-Stämmen T151023 und 12T272 frameshiftet.
Finegoldia magna bindet an Humanalbumin9. Ein Peptostreptokokken-Albumin-bindendes Protein (PAB) wurde im Stamm ALB8 untersucht; Seine Aktivität ist mit einer beschleunigten Wachstumsrate verbunden9. Das PAB-Protein des Stammes ALB8 (locus ID: X77864) enthält GA-Module, die Albumin-bindende Domäne; Die Struktur des GA-Moduls wurde aufgelöst19. Eine Blast-Suche ergab, dass ein enger PAB-Homolog in anderen sequenzierten Finegoldia-Genomen nicht gefunden werden kann (Tabelle S4B). Stattdessen gibt es mehrere Homologe, die eine Proteinidentität von bis zu 50% aufweisen; Ähnlichkeiten von Homologen in F. magna-Stämmen sind im Durchschnitt höher als bei Homologen in „F. nericia“ -Stämmen. In Bezug auf GAMMA-haltige Proteine sind insgesamt 16 Proteine in den 12 Finegoldia-Genomen kodiert, was auf eine gewisse Redundanz von albuminbindenden Proteinen hinweist (Tabelle S4C). Die meisten Stämme kodieren zwei Proteine mit GA-Modulen, mit Ausnahme von Stamm 09T408 (F. magna) und T151023 („F. nericia“).
Es wurde gezeigt, dass die an der Zellwand befestigte subtilisinähnliche Protease SufA mehrere menschliche Proteine wie Kollagen IV, Fibrinogen und antimikrobielle Peptide wie LL-37 und MIG / CXCL910 abbaut,20,21. Blast-Recherchen ergaben, dass die meisten Finegoldia-Stämme ein SufA-Homolog tragen, das dem untersuchten SufA aus Stamm ALB8 (Locus ID: DQ679960) sehr ähnlich ist (Proteinidentität > 75%). Ausnahmen sind die Stämme 07T609, 09T408 und CCUG54800. Ein zweites SufA-Homolog ist in den meisten Stämmen vorhanden; es hat eine durchschnittliche Proteinidentität von 26% (Tabelle S4D).
Ein weiterer wichtiger Wirt-wechselwirkender Faktor ist FAF (F. magna Adhäsionsfaktor). Etwa 90% der F. magna-Stämme produzieren diesen Faktor, der für die Verklumpung von Bakterien verantwortlich ist und die Bindung an die Basalmembran durch Bindung an BM-4011 vermittelt. Wir fanden heraus, dass sieben (drei F. magna- und vier „F. nericia“ -Stämme) der 12 Stämme ein FAF-Homolog codieren, das dem im ALB8-Stamm untersuchten ähnlich ist (Abb. 4B). Stammspezifische Unterschiede bestehen und Frameshift-Mutationen werden in den „F. nericia“ -Stämmen T151023 und 12T272 beobachtet. In den Stämmen 09T408, 09T494 und 12T306 findet sich kein Homolog (Tabelle S4E).
Finegoldia-Genome kodieren CAMP-Faktoren, die funktionell sind
Wir haben das Genom nach anderen mutmaßlichen Wirtsinteraktionsfaktoren durchsucht und Gene gefunden, die für Christie-Atkins-Munch-Petersen (CAMP) -Faktoren kodieren. CAMP-Faktoren können als Co-Hämolysine wirken und für die sogenannte CAMP-Reaktion verantwortlich sein, die synergistische Lyse von Schaferythrozyten durch Staphylococcus aureus Sphingomyelinase C (Beta-Toxin) und einen CAMP-Faktor22.
Genome Mining identifizierte zwei Gene, die CAMP-Faktoren in jedem genomsequenzierten Finegoldia-Stamm kodieren (CAMP1 und CAMP2; Abbildung S4); Jedes Homolog beherbergt die CAMP-Faktor-Familiendomäne (Pfam07373). Interessanterweise beherbergt ein Homolog (CAMP2) pro Stamm eine zusätzliche bakterielle Ig-ähnliche Domäne (Pfam02368). Neben CAMP1 und CAMP2 sind vier „F. nericia“ -Stämme enthalten zwei zusätzliche CAMP-Faktoren, die sich wesentlich von den anderen Homologen unterscheiden (CAMP3 und CAMP4; Abbildung S4). So beherbergen vier von sieben „F. nericia“ -Stämmen vier CAMP-Faktor-Gene.
Als nächstes wollten wir wissen, ob Finegoldia-Stämme eine positive CAMP-Reaktion hervorrufen. Ein CAMP-Agarplatten-Assay zeigt, dass einige Stämme stark CAMP-reaktionspositiv sind, während für andere Stämme unter den angewandten Kultivierungsbedingungen nach zwei und fünf Tagen Inkubation nur eine geringe Co-Hämolyse nachgewiesen werden kann (Abb. 5, Abbildung S5). Alle drei testeten F. magna-Stämme waren stark CAMP-reaktionspositiv. Unter den „F. nericia“ -Stämmen stellten wir eine gewisse Heterogenität fest; Zwei Stämme waren stark CAMP-reaktionspositiv, vier Stämme waren nur leicht co-hämolytisch und ein Stamm, CCUG54800, war eindeutig CAMP-reaktionspositiv. Die genomische Analyse ergab, dass im Stamm CCUG54800 das CAMP1-kodierende Gen eine Insertion eines Transposase-kodierenden Locus im 5′-Ende des Gens enthielt, der das Gen unterbricht (Abbildung S6).

CAMP-Reaktion von Finegoldia-Stämmen. Es wurden TSA-Agarplatten mit 5% Schafsblut verwendet. Staphylococcus aureus wird im mittleren Streifen beimpft. Ein positiver CAMP-Test wird durch vollständige Erythrozytenlyse an der Grenzfläche des Finegoldia sp. und die S. aureus-Streifen. Alle drei F. magna-Stämme (in Rot) und zwei von sieben „F. nericia“ -Stämmen (in blau) zeigten nach zwei Tagen anaerober Inkubation eine stark positive CAMP-Reaktion.
Unsere Ergebnisse zeigen, dass die Faktoren von Finegoldia sp. funktionell sein kann und dass es stammspezifische Unterschiede in der CAMP-Faktor-Aktivität gibt.
Zwei biochemische Testsysteme können F. magna nicht eindeutig von „F. nericia“ unterscheiden
Biochemische Reaktionen von GPAC einschließlich Stämmen von Finegoldia sp. wurden beschrieben; nur geringe biochemische Variation unter Finegoldia sp. dies wurde zuvor berichtet2,7. Wir haben zwei kommerziell erhältliche biochemische Testsysteme getestet, die zur Identifizierung von Anaerobiern verwendet werden. Das Kit „API® 20A“ enthält 21 Tests, darunter 16 Tests zur Kohlenhydratverwertung. Wie bereits berichtet, 7 konnte keines der Kohlenhydrate von Finegoldia sp. belastung (Daten nicht gezeigt). Die anderen Reaktionen (Indol, Urease, Katalase, Esculinhydrolyse und Gelatineverflüssigung) zeigten ebenfalls keinen Unterschied zwischen den Stämmen von F. magna und „F. nericia“ (Daten nicht gezeigt). Das Kit „RapIDTM ANA II“ enthält 18 Tests auf enzymatische Aktivitäten. Während saccharolytische Enzyme fehlen, sind alle Finegoldia sp. stämme produzieren proteolytische Enzyme (Tabelle S5). Die Reaktion zum Nachweis der Phosphataseaktivität war die einzige Reaktion, die sich zwischen den Stämmen unterschied, wobei alle F. magna-Stämme anscheinend negativ und die meisten „F. nericia“ -Stämme positiv waren. Die Interpretation dieses Testergebnisses ist jedoch begrenzt, da die Farbänderung an der Grenze der Signifikanz lag. Zusammenfassend lässt sich sagen, dass zwei bestehende kommerzielle biochemische Testsysteme, API® 20 A und RapIDTM ANA II, F. magna nicht eindeutig von „F. nericia“ unterscheiden können. Die biochemischen Eigenschaften von Finegoldia sp. müssen mit zusätzlichen biochemischen Tests weiter erforscht werden.
Mikroskopie zeigt Oberflächenunterschiede zwischen F. magna und „F. nericia“
In einem Versuch, die Morphologie und Oberflächenstruktur von F. magna und „F. nericia“ Wir verwendeten Rasterelektronenmikroskopie (SEM) und Rasterkraftmikroskopie (AFM) an Stämmen von F. magna (07T609, 08T492 und 09T408) und „F. nericia“ (09T494, 12T272 und 12T306). SEM erkannte Oberflächenunterschiede: Zellen von F. magna waren adhärenter oder aggregativer und produzierten scheinbar eine extrazelluläre Polymermatrix (Abb. 6). Dies bestätigt frühere Arbeiten, die eine Exopolysaccharidmatrix in Biofilmen von F. magna (Stamm FmBs12) durch Rasterelektronenmikroskopie sowie konfokale Laser-Scanning-Mikroskopie visualisiert haben23. Im getesteten „F. nericia“ Stämme konnten wir eine solche Matrix nicht identifizieren; Zellen scheinen weniger aggregativ zu sein.
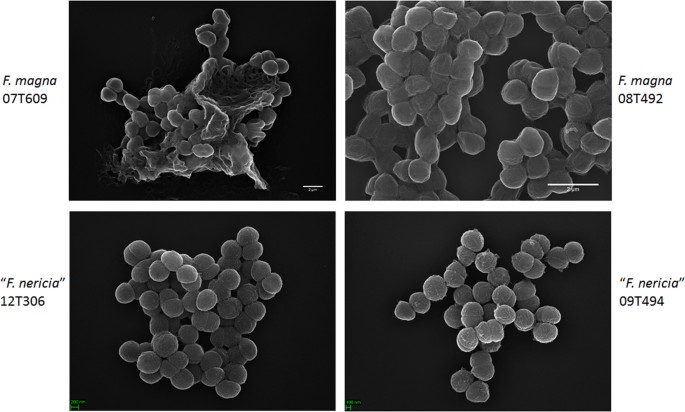
Rasterelektronenmikroskopischer Vergleich von F. magna und „F. nericia“. Die oberen und unteren Platten zeigen repräsentative Bilder von F. magna 07T609 und 08T492 und „F. nericia“ 09T494 und 12T306. Zellen von F. magna-Stämmen sind adhärenter / aggregativer und scheinen eine Polymermatrix zu erzeugen, insbesondere Stamm 07T609; Im Gegensatz dazu sind Zellen von „F. nericia“ weniger aggregativ.
AFM bestätigte diese Beobachtungen. Darüber hinaus beobachteten wir Unterschiede in Bezug auf Zellanhänge. Zellen von F. magna produzierten längere filamentöse Anhängsel, die aus der gesamten Zelloberfläche hervorstanden; Diese Strukturen könnten Pili oder Fimbrien darstellen (Abb. 7). Im Gegensatz dazu hatten Zellen von „F. nericia“ -Stämmen eine glattere Oberfläche ohne lange hervorstehende Anhängsel. Es kann jedoch nicht ausgeschlossen werden, dass diese AFM-Beobachtungen von den bakteriellen Kultivierungsbedingungen und von den Probenverarbeitungsschritten abhängen.

Rasterkraftmikroskopischer Vergleich von F. magna und „F. nericia“. Die linke und rechte Tafel stellen Bilder von F. magna 07T609 und „F. nericia“ 12T306 dar. (A und C) Hochauflösende Bilder mit der Advanced Soft-Touch-Methode mit Z-Skalenbalken bis 0,3 µm und XY-Balken bis 500 nm. (B und D) Überlagerungsbilder von kontrastreichen Bildern von (A und C), kombiniert mit unabhängiger horizontaler und vertikaler Polynomentfernung von (A und C), gelöst bis zum 11. Experimente wurden an drei Proben von jedem Stamm durchgeführt.