- Genomas de cepas de Finegoldia
- División de las cepas de Finegoldia en dos clados distintos basados en la comparación del genoma
- Heterogeneidad específica de la cepa de Finegoldia sp
- Factores que interactúan con el huésped y factores de virulencia (supuestos)
- Los genomas de Finegoldia codifican factores de CAMP que son funcionales
- Dos sistemas de pruebas bioquímicas no pueden diferenciar inequívocamente F. magna de»F. nericia»
- La microscopía revela diferencias superficiales entre F. magna y»F. nericia»
Genomas de cepas de Finegoldia
En total 17 genomas de Finegoldia sp. fueron analizados y comparados. Nuestro grupo15 ha secuenciado recientemente diez cepas, que se aislaron de pacientes con infecciones asociadas a implantes de articulaciones ortopédicas en Suecia. Siete genomas fueron secuenciados previamente por otros, incluyendo el genoma cerrado de la cepa tipo ATCC 2932814. Las características del genoma de todas las cepas de Finegoldia secuenciadas hasta ahora se resumen en la Tabla 1. La anotación con Prokka detectó entre 1570 y 1906 secuencias de codificación (CDS) por genoma, con un promedio de 1760 CDS por cepa.
División de las cepas de Finegoldia en dos clados distintos basados en la comparación del genoma
Anteriormente notamos una gran heterogeneidad entre los genomas de los aislados de Finegoldia 15. Se realizó una alineación del genoma central de los 17 genomas y se llamaron polimorfismos de nucleótido único (SNP) en el genoma central. En total, se identificaron 126.647 SNP del genoma central y se utilizaron para la reconstrucción filogenómica, revelando que los 17 genomas se pueden agrupar en dos clados distintos (Fig. 1). Un análisis adicional que determina el promedio de identificación de nucleótidos (ANI) del genoma central mostró que los dos clados exhiben un ANI del 90,7%, con una variación de ANI específica de la cepa del 90,2% al 91,2%. Un ANI tan bajo indica que los dos clados representan especies individuales16. Un clado contiene la mayoría de las cepas previamente secuenciadas y estudiadas, incluyendo ATCC29328 y ALB8; puede ser considerado como el clado clásico F. magna, con un ANI intraclade de 96,6% (variación 95,1–97,4%). El otro clado con un ANI intraclade del 94,8% (93,4-96.6%) contiene otra cepa ATCC (ATCC 53516) y principalmente cepas aisladas de pacientes del Hospital Universitario de Örebro, Suecia15. De aquí en adelante, las cepas de este clado se denominan tentativamente «Finegoldia nericia» (en latín, Närke, la región sueca, donde se aislaron la mayoría de las cepas de esta especie).

Árbol filogenómico basado en SNP del genoma central de los 17 genomas secuenciados de cepas de Finegoldia. El programa Parsnp se utilizó para alinear el genoma central y llamar SNPs. El genoma central está representado por el 43% del genoma de referencia (ATCC29328); se utilizaron un total de 126.647 SNP fiables del genoma central para reconstruir una filogenia del genoma completo. Los genomas se pueden asignar a dos clados principales; un clado representa cepas de F. magna y el otro clado consiste en cepas de una nueva especie, aquí llamada tentativamente «F. nericia».
Una comparación de secuencias de ARNr 16S de todos los aislados reveló un alto grado de similitud general (> 99%), pero detectó una separación de los aislados de F. magna de los aislados de «F. nericia», debido a 5 a 9 «F. SNP específicos de nericia (Figura S1). También reveló una mayor diversificación entre los aislados de «F. nericia» en comparación con F. magna. A continuación, analizamos secuencias adicionales de ARNr de 16S pertenecientes a Finegoldia sp. almacenado en GenBank (Figura S2). Esto reveló que la mayoría de las secuencias almacenadas se agrupan con «F. nericia», en particular un grupo de Finegoldia sp. cepas aisladas de biofilms de stent biliar17. Una vez más, se detecta una mayor diversificación entre los aislados de «F. nericia». A continuación, analizamos los datos del microbioma humano almacenados en la base de datos IMG (Genomas Microbianos integrados y muestras de microbioma) para detectar la presencia de secuencias de F. magna y «F. nericia». se pudieron identificar 3287 y 7068 secuencias codificantes de F. magna ATCC29328 y «F. nericia» ATCC 53516, respectivamente, en los 875 conjuntos de datos de microbioma humano ensamblados presentes en IMG (datos no mostrados); las ocurrencias más altas de Finegoldios se detectaron en muestras de pliegues retroauriculares. Tomados en conjunto, estos análisis sugieren que entre los aislados de Finegoldios asociados con humanos, «F. nericia» parece ser más prevalente que F. magna.
Heterogeneidad específica de la cepa de Finegoldia sp
Para arrojar más luz sobre la estructura de la población del género Finegoldia, se llevaron a cabo análisis comparativos de 12 genomas (las diez cepas recién secuenciadas y las dos cepas ATCC), incluidas cuatro cepas del clado F. magna y ocho cepas del clado «F. nericia». Una comparación visual de los 12 genomas resaltó la diferencia de identidad de nucleótidos entre los dos clados (Fig. 2). Curiosamente, el plásmido encontrado en ATCC 29328 parece ser específico para esta cepa, ya que ninguna otra cepa secuenciada contiene una secuencia similar (Fig. 2A).

comparación del Genoma de 12 cepas pertenecientes al género Finegoldia. Los dos anillos más internos representan el contenido de G + C (negro) y el sesgo de GC (violeta/verde). (A) La cepa de referencia es ATCC29328 (F. magna); nótese la alta identidad de nucleótidos (visualizada por el color rojo oscuro) de los tres genomas más internos (pertenecientes todos a F. magna), y la inferior se identifica con ocho cepas pertenecientes a «F. nericia» (color rojo pálido de los ocho anillos exteriores); de adentro hacia afuera: cepas 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 y ATCC 53516. La gran región específica ATCC29328 en la región superior izquierda representa el plásmido pFMC. B) La cepa de referencia es 09T494 («F. nericia»). Los siete genomas de «F. nericia» (anillos interiores) tienen un nucleótido más alto (representado por el color rojo más oscuro), en comparación con los cuatro genomas de F. magna (anillos exteriores); de adentro hacia afuera: cepas 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 y 09T408. No hay signos de regiones genómicas específicas de «F. nericia», pero tenga en cuenta los cinco grandes grupos específicos de cepas.
No se detectaron diferencias significativas en el tamaño del genoma y el número de CDS entre los dos clados. Además, no se detectaron regiones genómicas específicas de clado (es decir, específicas de F. magna o «F. nericia»), según se juzgó a partir del análisis de BRIG (Fig. 2) y también confirmado por un enfoque blast bidireccional de todos los CD utilizando ProteinOrtho (Tabla S1). Este enfoque detectó 1202 ortólogos compartidos por los 12 genomas, es decir, en promedio el 68% de los CD de cada cepa son parte del proteoma central. Solo se encuentran muy pocos genes específicos de clado. En cambio, se pueden encontrar muchas regiones compartidas por solo unas pocas cepas, por ejemplo, para la cepa 09T494 (véase también la Fig. 2B). Además, existen regiones específicas de cepa que codifican un total de 1016 CDS específicos de cepa, con un amplio rango entre las cepas, por ejemplo, se detectan 187 y 22 CDS específicos de cepa en la cepa ATCC29328 y T151023, respectivamente (Tabla S1). Entre las funciones específicas de la cepa se encuentran varias funciones de transporte (hierro, bombas de eflujo, transportadores de oligopéptidos ABC), proteínas de superficie, sistemas de restricción y modificación, funciones de transferencia conjugativa, síntesis de poliquétidos, bacteriocinas, funciones relacionadas con fagos, determinantes de la resistencia a los antibióticos y sistemas CRISPR/cas.
Curiosamente, se detectaron diferencias en los sistemas CRISPR/cas entre cepas de F. magna y «F. nericia» (Tabla S2). Siete de nueve F. las cepas magna contenían de uno a cuatro matrices CRISPR por cepa, conteniendo en promedio 12 espaciadores (que oscilaban entre 3 y 38 espaciadores por cepa). La repetición «GTTTGAGAATGATGTAATTTCATATAGGTATTAAAC» fue específica de cepas de F. magna. En contraste ,las cepas de «F. nericia» llevaban solo una matriz CRISPR por cepa que contenía un promedio de 53 espaciadores (rango de 14 a 87 por cepa). Cada cepa contenía un conjunto individual de espaciadores en su(s) matriz (s) CRISPR, lo que indica que cada cepa tiene una historia evolutiva única.
Para evaluar si tales funciones específicas de deformación se adquirieron horizontalmente, observamos signos de su movilidad/adquisición utilizando el programa IslandViewer (Figura S3). Este programa predijo de 141 (cepa 09T408) a 408 (cepa 12T273) genes ubicados en islas genómicas que es probable que se adquieran horizontalmente, incluidos muchos de los genes específicos de la cepa mencionados anteriormente que participan en la especialización, la aptitud, la supervivencia y la competencia entre especies (Tabla S3).
Factores que interactúan con el huésped y factores de virulencia (supuestos)
Se han descrito varios factores que interactúan con el huésped para F. magna 4,5. Se han identificado pili dependientes de sortasa que son importantes para la adhesión y la colonización13. Se conserva un locus genómico que codifica los genes de la subunidad pilus Fmp1, la subunidad putativa Fmp2 y tres sortasas entre todos los genomas secuenciados (Fig. 3A). Hay variaciones específicas de la cepa arriba y abajo de este locus, i. e. algunas cepas poseen genes adicionales aguas arriba de fmp1 y aguas abajo del locus de la sortasa, incluyendo un gen que codifica una proteína de la familia hemolisina III. Curiosamente, hay diferencias específicas de cepa con respecto a fmp1. Casi cada cepa lleva una variante individual y un análisis filogenético de Fmp1 a nivel de proteína no mostró correlación con el análisis filogenómico del genoma central (Fig. 3B). Por el contrario, la Fmp2 está altamente conservada entre todas las cepas. Otra proteína, codificada aguas arriba de fmp1, alberga dominios de unión de colágeno y fibrinógeno; el dominio de unión de colágeno (COG4932) también está presente en Fmp2. Proponemos que esta proteína también pertenezca al locus pilus y llamemos al gen correspondiente fmp3.

Loci genómicos codificadores pili dependientes de sortasa en los genomas de cepas de Finegoldia. A) Organización genómica de los loci de las cepas de F. magna (rojo) y «F. nericia» (azul claro). Los tres genes de la sortasa se representan en verde claro (etiquetados como «2»); el gen de la subunidad pilus mayor Fmp1 se muestra en rojo (marcado con «1») y fmp2 se muestra en naranja (marcado con»3″). Aguas arriba de fmp1 y aguas abajo de los genes de la sortasa en algunas cepas hay genes adicionales (etiquetados como «8», proteína de la familia hemolisina III y «10», proteína hipotética) presentes. Una supuesta proteína de unión al colágeno, Fmp3 (etiquetada como «4»), está codificada aguas arriba de fmp1. B) Árbol filogenético de Fmp1 de cepas de Finegoldia. La mayoría de las cepas poseen una variante Fmp1 individual. La historia evolutiva se infirió utilizando el método de Máxima Verosimilitud, realizado en MEGA7.
La proteína L es probablemente el factor mejor estudiado de F. magna 8. Es un superantígeno que se dirige a las células B y a las inmunoglobulinas. Se ha detectado en aproximadamente el 10% de los aislados de Finegoldia hasta ahora probados, pero varía en tamaño y número de dominios de unión de fragmentos de unión a antígenos (Fab) (dominio B1, pfam02246)4,8,18. Una búsqueda explosiva con la proteína L estudiada de la cepa 312 (locus ID: M86697, 719 aa) revela algunas proteínas muy similares (identidad de proteína superior al 50%) con tamaños variables (603 a 1417 aa) en unas pocas cepas, p.ej. cepas CCUG54800, T161024, ATCC53516, 12T273 y 12T306 (Fig. 4A). Todas estas son cepas de «F. nericia», lo que indica que el superantígeno de células B es más prevalente en esta especie. La proteína excepcionalmente grande (1417 aa) factor tipo L en la cepa 12T306 posee además de dos dominios B1 también dos módulos GA que se conocen a partir de proteínas peptostreptocócicas de unión a albúmina (ver a continuación). Muchas proteínas adicionales con baja similitud a la proteína L, pero sin contener dominios B1 aparentes, están codificadas en las cepas de Finegoldia secuenciadas por el genoma (Tabla S4A); una vez más, existen variaciones específicas de la cepa, y algunas cepas, es decir, 09T408, 09T494 y T151023, no poseen tales homólogos.

Loci genómicos que codifican a los homólogos de proteínas L y FAF en cepas de Finegoldia. A) En cepas Finegoldia secuenciadas genómicamente se realizaron búsquedas blandas con la proteína L caracterizada de la cepa 312 (locus ID: M86697) y se muestran homólogos (en rojo, etiquetados con «1») con alta similitud (>50% de identidad proteica) (véase también el cuadro S4A). Solo cepas de» F. nericia», pero no de F. las cepas magna contienen tales homólogos con longitudes variables. B) Las búsquedas por explosión con la proteína FAF estudiada de la cepa ALB8 encontraron homólogos de longitud completa (en rojo, etiquetados con «1») en tres cepas de F. magna (roja) y cuatro de «F. nericia» (azul claro). El gen FAF se enmarca en las dos cepas de» F. nericia » T151023 y 12T272.
Se encontró que Finegoldia magna se une a la albúmina humana9. Se ha estudiado una proteína peptostreptocócica de unión a albúmina (PAB) en la cepa ALB8; su actividad está relacionada con una tasa de crecimiento acelerado9. La proteína PAB de la cepa ALB8 (locus ID: X77864) contiene módulos GA, el dominio de unión a la albúmina; se ha resuelto la estructura del módulo GA19. Una búsqueda por Explosión reveló que no se puede encontrar un homólogo de PAB cercano en otros genomas Finegoldios secuenciados (Tabla S4B). En cambio, hay varios homólogos que comparten hasta un 50% de identidad proteica; las similitudes de los homólogos en las cepas de F. magna son en promedio más altas en comparación con los homólogos en las cepas de «F. nericia». En cuanto a las proteínas que contienen módulos GA, en total 16 proteínas están codificadas en los 12 genomas de Finegoldia, lo que indica cierta redundancia de proteínas de unión a albúmina (Tabla S4C). La mayoría de las cepas codifican dos proteínas con módulos GA, excepto las cepas 09T408 (F. magna) y T151023 («F. nericia»).
Se ha demostrado que la proteasa SufA subtilisínica unida a la pared celular degrada varias proteínas humanas, como colágeno IV, fibrinógeno y péptidos antimicrobianos como LL-37 y MIG/CXCL910,20,21. Las búsquedas por explosión determinaron que la mayoría de las cepas de Finegoldia tienen un homólogo de SufA que es muy similar (identidad de proteína > 75%) al SufA estudiado de la cepa ALB8 (locus ID: DQ679960). Las excepciones son las cepas 07T609, 09T408 y CCUG54800. Un segundo homólogo SufA está presente en la mayoría de las cepas; tiene una identidad proteica promedio del 26% (Tabla S4D).
Otro factor importante que interactúa con el huésped es el FAF (factor de adhesión F. magna). Alrededor del 90% de las cepas de F. magna producen este factor, que es responsable de la acumulación de bacterias y media la unión a la membrana basal al unirse a BM-4011. Encontramos que siete (tres cepas de F. magna y cuatro de «F. nericia») de las 12 cepas codifican un homólogo FAF similar al estudiado en la cepa ALB8 (Fig. 4B). Existen diferencias específicas de la cepa y se observan mutaciones de cambio de marco en las cepas T151023 y 12T272 de «F. nericia». No se puede encontrar homólogo en las cepas 09T408, 09T494 y 12T306 (Tabla S4E).
Los genomas de Finegoldia codifican factores de CAMP que son funcionales
Buscamos en el genoma otros factores putativos que interactúan con el huésped y encontramos genes que codifican factores de Christie-Atkins-Munch-Petersen (CAMP). Los factores CAMP pueden actuar como co-hemolisinas y dar cuenta de la llamada reacción CAMP, la lisis sinérgica de eritrocitos ovinos por la esfingomielinasa C (toxina beta) de Staphylococcus aureus y un factor camp22.
La minería genómica identificó dos genes que codifican factores CAMP en cada cepa Finegoldia secuenciada por genoma (CAMP1 y CAMP2; Figura S4); cada homólogo alberga el dominio de la familia de factores CAMP (Pfam07373). Curiosamente, un homólogo (CAMP2) por cepa alberga un dominio bacteriano similar a Ig adicional (Pfam02368). Además de CAMP1 y CAMP2, cuatro » F. las cepas de nericia contienen dos factores CAMP adicionales que difieren sustancialmente de los otros homólogos (CAMP3 y CAMP4; Figura S4). Por lo tanto, cuatro de cada siete cepas de «F. nericia» albergan cuatro genes del factor CAMP.
A continuación, queríamos saber si las cepas de Finegoldia producen una reacción positiva al CAMP. Un ensayo en placa de agar CAMP muestra que algunas cepas son fuertemente positivas a la reacción CAMP, mientras que para otras cepas solo se puede detectar una pequeña co-hemólisis en las condiciones de cultivo aplicadas a los dos y cinco días de incubación (Fig. 5, figura S5). Los tres probaron F. las cepas magna fueron fuertemente positivas a la reacción CAMP. Entre las cepas de» F. nericia » detectamos cierta heterogeneidad; dos cepas fueron fuertemente positivas a la reacción CAMP, cuatro cepas fueron solo ligeramente co-hemolíticas y una cepa, CCUG54800, fue claramente negativa a la reacción CAMP. El análisis genómico encontró que en la cepa CCUG54800 el gen codificador de CAMP1 contenía una inserción de un locus codificador de transposasa en el extremo 5’del gen, que altera el gen (Figura S6).

Reacción CAMP de cepas de Finegoldia. Se utilizaron placas de agar TSA con un 5% de sangre de oveja. Staphylococcus aureus está inoculado en la raya media. Una prueba de AMPC positiva se indica por lisis eritrocitaria completa en la interfaz de Finegoldia sp. y las rayas de S. Aureus. Las tres cepas de F. magna (en rojo) y dos de las siete cepas de «F. nericia» (en azul) mostraron una fuerte reacción positiva de CAMP después de dos días de incubación anaeróbica.
Nuestros resultados indican que los factores CAMP de Finegoldia sp. puede ser funcional y que hay diferencias específicas de la cepa en la actividad del factor CAMPAMENTO.
Dos sistemas de pruebas bioquímicas no pueden diferenciar inequívocamente F. magna de»F. nericia»
Reacciones bioquímicas de GPAC, incluidas cepas de Finegoldia sp. se han descrito; solo poca variación bioquímica entre Finegoldia sp. las cepas fueron previamente notificadas2, 7. Probamos dos sistemas de pruebas bioquímicas disponibles en el mercado que se utilizan para la identificación de anaerobios. El kit «API® 20A» contiene 21 pruebas, entre ellas 16 pruebas para la utilización de carbohidratos. Como se ha informado previamente7, ninguno de los hidratos de carbono pudo ser metabolizado por ningún Finegoldia sp. cepa (no se muestran los datos). Las otras reacciones (indol, ureasa, catalasa, hidrólisis de esculina y licuefacción de gelatina) tampoco revelaron ninguna diferencia entre las cepas de F. magna y «F. nericia» (datos no mostrados). El kit «RapIDTM ANA II» contiene 18 pruebas para actividades enzimáticas. Mientras que faltan enzimas sacarolíticas, todas las Finegoldia sp. las cepas producen enzimas proteolíticas (Tabla S5). La reacción que detectó la actividad de la fosfatasa fue la única reacción que difería entre las cepas, con todas las cepas de F. magna aparentemente negativas y la mayoría de las cepas de «F. nericia» positivas. Sin embargo, la interpretación de este resultado de la prueba es limitada, ya que el cambio de color estaba en el límite de significación. En conclusión, dos sistemas de pruebas bioquímicas comerciales existentes, API ® 20 A y RapIDTM ANA II, no pueden diferenciar inequívocamente a F. magna de «F. nericia». Las propiedades bioquímicas de Finegoldia sp. deben explorarse más a fondo con pruebas bioquímicas adicionales.
La microscopía revela diferencias superficiales entre F. magna y»F. nericia»
En un intento de visualizar la morfología y la estructura superficial de F. magna y » F. nericia » utilizamos microscopía electrónica de barrido (SEM) y microscopía de fuerza atómica (AFM) en cepas de F. magna (07T609, 08T492 y 09T408) y «F. nericia» (09T494, 12T272 y 12T306). El SEM detectó diferencias superficiales: las células de F. magna eran más adherentes o agregativas, aparentemente produciendo una matriz polimérica extracelular (Fig. 6). Esto confirma trabajos previos que han visualizado una matriz de exopolisacáridos en biopelículas de F. magna (cepa FmBs12) mediante microscopía electrónica de barrido, así como microscopía de barrido láser confocal23. En la probada » F. nericia » cepas no pudimos identificar tal matriz; las células parecen menos agregativas.
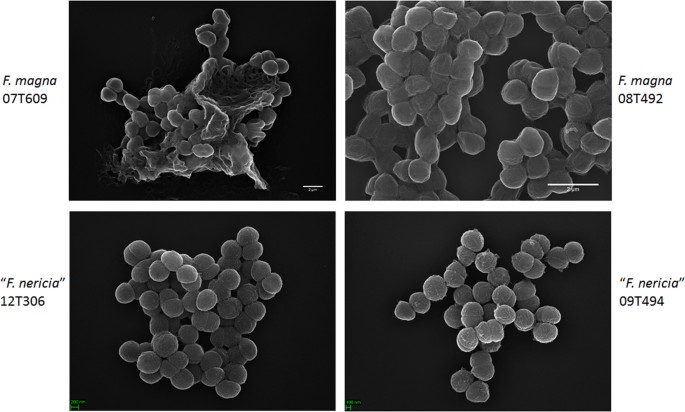
Comparación microscópica electrónica de barrido de F. magna y «F. nericia». Los paneles superior e inferior muestran imágenes representativas de F. magna 07T609 y 08T492 y «F. nericia» 09T494 y 12T306, respectivamente. Las células de las cepas de F. magna son más adherentes / agregativas y parecen producir una matriz polimérica, en particular la cepa 07T609; en contraste, las células de «F. nericia» son menos agregativas.
AFM confirmaron estas observaciones. Además, se observaron diferencias con respecto a los apéndices celulares. Las células de F. magna produjeron apéndices filamentosos más largos que sobresalían de toda la superficie celular; estas estructuras podrían representar pili o fimbrias (Fig. 7). En contraste, las células de las cepas de «F. nericia» tenían una superficie más lisa sin apéndices salientes largos. Sin embargo, no se puede descartar que estas observaciones de AFM dependan de las condiciones de cultivo bacteriano y de las etapas de procesamiento de las muestras.

Comparación microscópica de fuerza atómica de F. magna y «F. nericia». Los paneles izquierdo y derecho representan imágenes de F. magna 07T609 y» F. nericia » 12T306, respectivamente. (A y C) Imágenes de alta resolución con método avanzado asistido de tacto suave con barras de escala Z de hasta 0,3 µm y barras XY que indican 500 nm. (B y D) Imágenes superpuestas de imágenes de alto contraste de (A y C), combinadas con eliminación de polinomios horizontales y verticales independientes de (A y C), resueltas hasta el grado 11. Se realizaron experimentos con tres muestras de cada cepa.