- Genomi di ceppi di Finegoldia
- Divisione dei ceppi di Finegoldia in due cladi distinti basati sul confronto del genoma
- Eterogeneità ceppo-specifica di Finegoldia sp
- Fattori di interazione con l’ospite e fattori di virulenza (putativi)
- I genomi di Finegoldia codificano i fattori di CAMP funzionali
- Due sistemi di test biochimici non possono distinguere univocamente F. magna da”F. nericia”
- La microscopia rivela differenze superficiali tra F. magna e”F. nericia”
Genomi di ceppi di Finegoldia
In totale 17 genomi di Finegoldia sp. sono stati analizzati e confrontati. Dieci ceppi sono stati recentemente sequenziati dal nostro gruppo15; sono stati isolati da pazienti con infezioni associate a protesi articolari ortopediche in Svezia. Sette genomi sono stati precedentemente sequenziati da altri, incluso il genoma chiuso del ceppo tipo ATCC 2932814. Le caratteristiche del genoma di tutti i ceppi finora sequenziati di Finegoldia sono riassunte nella Tabella 1. L’annotazione utilizzando Prokka rilevato tra 1570 e 1906 sequenze di codifica (CDS) per genoma, con una media di 1760 CDS per ceppo.
Divisione dei ceppi di Finegoldia in due cladi distinti basati sul confronto del genoma
In precedenza abbiamo notato una grande eterogeneità tra i genomi degli isolati di Finegoldia15. È stato fatto un allineamento core-genoma di tutti i genomi 17 e sono stati chiamati polimorfismi a singolo nucleotide (SNPS) nel genoma principale. In totale, 126.647 core-genome SNPs sono stati identificati e utilizzati per la ricostruzione filogenomica, rivelando che i 17 genomi possono essere raggruppati in due cladi distinti (Fig. 1). Un’analisi supplementare che determina il nucleotide medio identifica (ANI) del genoma del centro ha indicato che i due cladi esibiscono un ANI di 90,7%, con una variazione ANI ceppo-specifica da 90,2% a 91,2%. Un ANI così basso indica che i due cladi rappresentano singole specie16. Un clade contiene la maggior parte dei ceppi precedentemente sequenziati e studiati, tra cui ATCC29328 e ALB8; può essere considerato come il classico clade F. magna, con un ANI intraclade del 96,6% (variazione 95,1–97,4%). L’altro clade con un ANI intraclade del 94,8% (93,4–96.6%) contiene un altro ceppo ATCC (ATCC 53516) e principalmente ceppi isolati da pazienti dell’Ospedale universitario di Öebro, in Svezia15. In seguito, i ceppi di questo clade sono provvisoriamente chiamati “Finegoldia nericia” (latino per Närke, la regione svedese, dove la maggior parte dei ceppi di questa specie sono stati isolati).

Albero filogenomico basato sul nucleo-genoma SNPS di tutti i 17 genomi sequenziati di ceppi Finegoldia. Il programma Pastinaca è stato utilizzato per allineare il genoma del nucleo e chiamare SNPS. Il genoma di base è rappresentato dal 43% del genoma di riferimento (ATCC29328); un totale di 126.647 SNPS nucleo-genoma affidabili sono stati utilizzati per ricostruire un intero genoma filogenesi. I genomi possono essere assegnati a due cladi principali; un clade rappresenta i ceppi di F. magna e l’altro clade è costituito da ceppi di una nuova specie, qui provvisoriamente chiamata “F. nericia”.
Un confronto di sequenze di rRNA 16S di tutti gli isolati ha rivelato un alto grado di somiglianza complessiva (>99%), ma ha rilevato una separazione di F. magna isolati da “F. nericia” isolati, a causa di 5 a 9 “F. nericia ” – SNP specifici (Figura S1). Ha anche rivelato una maggiore diversificazione tra gli isolati di “F. nericia” rispetto a F. magna. Successivamente, abbiamo analizzato ulteriori sequenze rRNA 16S appartenenti a Finegoldia sp. memorizzato in GenBank (Figura S2). Ciò ha rivelato che la maggior parte delle sequenze memorizzate si raggruppano con “F. nericia”, in particolare un gruppo di Finegoldia sp. ceppi isolati da biofilms17 stent biliare. Ancora una volta, viene rilevata una maggiore diversificazione tra gli isolati di “F. nericia”. Successivamente, abbiamo analizzato i dati del microbioma umano memorizzati nel database IMG (Integrated Microbical Genomes and microbiome samples) per la presenza di sequenze di F. magna e “F. nericia”. 3287 e 7068 sequenze codificanti di F. magna ATCC29328 e” F. nericia ” ATCC 53516, rispettivamente, potrebbero essere identificate negli 875 set di dati del microbioma umano assemblati presenti in IMG( dati non mostrati); le più alte occorrenze di Finegoldia sono state rilevate in campioni di piega retroauricolare. Nel loro insieme, queste analisi suggeriscono che tra gli isolati di Finegoldia associati all’uomo “F. nericia” sembra essere più prevalente di F. magna.
Eterogeneità ceppo-specifica di Finegoldia sp
Per fare ulteriore luce sulla struttura della popolazione del genere Finegoldia, sono state effettuate analisi comparative di 12 genomi (tutti i dieci ceppi appena sequenziati e i due ceppi ATCC), inclusi quattro ceppi del clade F. magna e otto ceppi del clade “F. nericia”. Un confronto visivo dei 12 genomi ha evidenziato la differenza di identità nucleotidica tra i due cladi (Fig. 2). È interessante notare che il plasmide trovato in ATCC 29328 sembra essere specifico per questo ceppo in quanto nessun altro ceppo sequenziato contiene una sequenza simile (Fig. 2 BIS).

Confronto genoma di 12 ceppi appartenenti al genere Finegoldia. I due anelli più interni rappresentano il G + C-content (nero) e il GC-skew (viola/verde). (A) Il ceppo di riferimento è ATCC29328 (F. magna); notare l’alta identità nucleotidica (visualizzata dal colore rosso scuro) dei tre genomi più interni (appartenenti tutti a F. magna), e il più basso identificano otto ceppi appartenenti a “F. nericia” (colore rosso pallido degli otto anelli esterni); dall’interno all’esterno: ceppi 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 e ATCC 53516. La grande regione ATCC29328-specifica nella regione in alto a sinistra rappresenta il plasmide pFMC. B) Il ceppo di riferimento è 09T494 (“F. nericia”). I sette genomi “F. nericia” (anelli interni) hanno un nucleotide più elevato (rappresentato dal colore rosso più scuro), rispetto ai quattro genomi F. magna (anelli esterni); dall’interno verso l’esterno: ceppi 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 e 09T408. Non ci sono segni di regioni genomiche specifiche di “F. nericia”, ma si noti i cinque grandi cluster specifici del ceppo.
Non sono state rilevate differenze significative nella dimensione del genoma e nel numero di CD tra i due cladi. Inoltre, non sono state rilevate regioni genomiche specifiche del clade (cioè F. magna o “F. nericia”), come giudicato dall’analisi BRIG (Fig. 2) e confermato anche da un approccio blast bidirezionale di tutti i CD utilizzando ProteinOrtho (Tabella S1). Questo approccio ha rilevato 1202 ortologhi condivisi da tutti i 12 genomi, cioè in media il 68% dei CDS di ciascun ceppo fa parte del proteoma principale. Solo pochissimi geni clade-specifici sono stati trovati. Invece, si possono trovare molte regioni condivise da solo pochi ceppi, ad esempio per il ceppo 09T494 (vedi anche Fig. 2 TER). Inoltre, sono presenti regioni specifiche del ceppo che codificano in totale 1016 CD specifici del ceppo, con un ampio intervallo tra i ceppi, ad esempio 187 e 22 CD specifici del ceppo sono rilevati rispettivamente nel ceppo ATCC29328 e T151023 (Tabella S1). Tra le funzioni specifiche del ceppo ci sono varie funzioni di trasporto (ferro, pompe di efflusso, trasportatori oligopeptidi ABC), proteine di superficie, sistemi di restrizione-modifica, funzioni di trasferimento coniugativo, sintesi di polichetidi, batteriocine, funzioni correlate ai fagi, determinanti della resistenza agli antibiotici e sistemi CRISPR/cas.
È interessante notare che sono state rilevate differenze nei sistemi CRISPR/cas tra ceppi di F. magna e “F. nericia” (Tabella S2). Sette su nove F. i ceppi magna contenevano da uno a quattro array CRISPR per ceppo, contenenti in media 12 distanziatori (da 3 a 38 distanziatori per ceppo). La ripetizione “GTTTGAGAATGATGATGTAATTTCATATAGGTATTAAAC” era specifica per i ceppi di F. magna. Al contrario, i ceppi “F. nericia” trasportavano solo un array CRISPR per ceppo che conteneva in media 53 distanziatori (intervallo da 14 a 87 per ceppo). Ogni ceppo conteneva un set individuale di distanziatori nei loro array CRISPR, sottendendo l’eterogeneità specifica del ceppo e indicando che ogni ceppo ha una storia evolutiva unica.
Per valutare se tali funzioni specifiche del ceppo sono state acquisite orizzontalmente, abbiamo esaminato i segni della loro mobilità/acquisizione utilizzando il programma IslandViewer (Figura S3). Questo programma prevedeva da 141 (ceppo 09T408) a 408 (ceppo 12T273) geni situati in isole genomiche che potrebbero essere acquisite orizzontalmente, inclusi molti dei geni specifici del ceppo sopra menzionati coinvolti nella specializzazione, fitness, sopravvivenza e competizione interspecie (Tabella S3).
Fattori di interazione con l’ospite e fattori di virulenza (putativi)
Diversi fattori di interazione con l’ospite sono stati descritti per F. magna 4,5. Sono stati identificati pili Sortase-dipendenti che sono importanti per l’adesione e la colonizzazione13. Un locus genomico che codifica i geni per la subunità pilus Fmp1, la subunità putativa Fmp2 e tre sortasi è conservato tra tutti i genomi sequenziati (Fig. 3 BIS). Ci sono variazioni specifiche del ceppo a monte e a valle di questo locus, cioè alcuni ceppi possiedono geni aggiuntivi a monte di fmp1 e a valle del locus sortasi, tra cui un gene che codifica una proteina della famiglia emolisina III. È interessante notare che ci sono differenze specifiche del ceppo per quanto riguarda fmp1. Quasi ogni ceppo porta una variante individuale e un’analisi filogenetica di Fmp1 a livello proteico non ha mostrato alcuna correlazione con l’analisi filogenomica del genoma del nucleo (Fig. 3 TER). Al contrario, Fmp2 è altamente conservato tra tutti i ceppi. Un’altra proteina, codificata a monte di fmp1, ospita i domini di legame del collagene e del fibrinogeno; il dominio di legame del collagene (COG4932) è presente anche in Fmp2. Proponiamo che questa proteina appartenga anche al locus pilus e chiami il gene corrispondente fmp3.

Loci genomici codificanti pili-dipendenti da Sortasi nei genomi dei ceppi di Finegoldia. (A) Organizzazione genomica dei loci nei ceppi F. magna (rosso) e “F. nericia” (azzurro). I tre geni sortasi sono raffigurati in verde chiaro (etichettato “2”); il gene per la subunità pilus maggiore Fmp1 è mostrato in rosso (etichettato “1”) e fmp2 è raffigurato in arancione (etichettato”3″). A monte di fmp1 e a valle dei geni sortasi in alcuni ceppi sono presenti geni aggiuntivi (etichettati “8”, proteina della famiglia dell’emolisina III e “10”, proteina ipotetica). Una presunta proteina legante il collagene, Fmp3 (etichettata “4”), è codificata a monte di fmp1. (B) Albero filogenetico di Fmp1 di ceppi Finegoldia. La maggior parte dei ceppi possiede una variante Fmp1 individuale. La storia evolutiva è stata dedotta utilizzando il metodo di massima verosimiglianza, fatto in MEGA7.
La proteina L è probabilmente il fattore meglio studiato di F. magna 8. È un superantigene che mira alle cellule B e alle immunoglobuline. È stato rilevato in circa il 10% degli isolati Finegoldia finora testati, ma varia in termini di dimensioni e numero di domini di legame del frammento di antigene (Fab) (dominio B1, pfam02246)4,8,18. Una ricerca blast con la proteina L studiata dal ceppo 312 (locus ID: M86697, 719 aa) rivela alcune proteine altamente simili (identità proteica superiore al 50%) con dimensioni variabili (da 603 a 1417 aa) in alcuni ceppi, cioè ceppi CCUG54800, T161024, ATCC53516, 12T273 e 12T306 (Fig. 4 BIS). Tutti questi sono ceppi “F. nericia”, il che indica che il superantigene a cellule B è più diffuso in questa specie. Il fattore L-like della proteina eccezionalmente grande (1417 aa) nel ceppo 12T306 possiede oltre a due domini B1 anche due moduli GA che sono noti dalle proteine peptostreptococciche che legano l’albumina (vedi sotto). Molte proteine aggiuntive con bassa somiglianza con la proteina L, ma senza contenere alcun dominio B1 apparente sono codificate nei ceppi Finegoldia sequenziati dal genoma (Tabella S4A); ancora una volta, esistono variazioni specifiche del ceppo e alcuni ceppi, cioè 09T408, 09T494 e T151023 non possiedono tali omologhi.

Loci genomici che codificano gli omologhi della proteina L e FAF nei ceppi di Finegoldia. (A) Ricerche blast con la proteina L caratterizzata dal ceppo 312 (locus ID: M86697) sono state effettuate in ceppi di Finegoldia sequenziati dal genoma e sono mostrati omologhi (in rosso, etichettati “1”) con elevata somiglianza (>50% di identità proteica) (vedi anche Tabella S4A). Solo ceppi “F. nericia” ma non F. i ceppi magna contengono tali omologhi con lunghezze variabili. (B) Ricerche blast con la proteina FAF studiata del ceppo ALB8 hanno trovato omologhi a lunghezza intera (in rosso, etichettati “1”) in tre ceppi F. magna (rosso) e quattro “F. nericia” (azzurro). Il gene FAF è frameshifted nei due ceppi “F. nericia” T151023 e 12T272.
Finegoldia magna è stato trovato per legare all’albumina umana9. Nel ceppo ALB8 è stata studiata una proteina peptostreptococcica che lega l’albumina (PAB); la sua attività è legata ad un tasso di crescita accelerato9. La proteina PAB del ceppo ALB8 (locus ID: X77864) contiene moduli GA, il dominio di legame all’albumina; la struttura del modulo GA è stata risoluta19. Una ricerca Blast ha rivelato che un omologo PAB vicino non può essere trovato in altri genomi Finegoldia sequenziati (Tabella S4B). Invece, ci sono diversi omologhi che condividono fino al 50% di identità proteica; le somiglianze degli omologhi nei ceppi di F. magna sono in media più alte rispetto agli omologhi nei ceppi di “F. nericia”. Per quanto riguarda le proteine contenenti moduli GA, in totale le proteine 16 sono codificate nei genomi 12 Finegoldia, indicando una certa ridondanza delle proteine leganti l’albumina (Tabella S4C). La maggior parte dei ceppi codificano due proteine con moduli GA, ad eccezione del ceppo 09T408 (F. magna) e T151023 (“F. nericia”).
La proteasi simile alla subtilisina attaccata alla parete cellulare SufA ha dimostrato di degradare diverse proteine umane, come il collagene IV, il fibrinogeno e i peptidi antimicrobici come LL-37 e MIG/CXCL910,20,21. Le ricerche blast hanno determinato che la maggior parte dei ceppi di Finegoldia ha un omologo SufA altamente simile (identità proteica > 75%) al SufA studiato dal ceppo ALB8 (locus ID: DQ679960). Le eccezioni sono i ceppi 07T609, 09T408 e CCUG54800. Un secondo omologo SufA è presente nella maggior parte dei ceppi; ha un’identità proteica media del 26% (Tabella S4D).
Un altro importante fattore di interazione ospite è il FAF (F. magna adhesion factor). Circa il 90% dei ceppi di F. magna produce questo fattore, che è responsabile dell’aggregazione dei batteri e media il legame con la membrana basale legandosi a BM-4011. Abbiamo scoperto che sette (tre F. magna e quattro ceppi “F. nericia”) dei 12 ceppi codificano un omologo FAF simile a quello studiato nel ceppo ALB8 (Fig. 4 TER). Esistono differenze specifiche del ceppo e mutazioni di frameshift sono osservate nei ceppi “F. nericia” T151023 e 12T272. Nessun omologo può essere trovato nei ceppi 09T408, 09T494 e 12T306 (Tabella S4E).
I genomi di Finegoldia codificano i fattori di CAMP funzionali
Abbiamo cercato nel genoma altri fattori che interagiscono con l’ospite e abbiamo trovato geni che codificano per i fattori di Christie-Atkins-Munch-Petersen (CAMP). I fattori CAMP possono agire come co-emolisine e rappresentano la cosiddetta reazione CAMP, la lisi sinergica degli eritrociti di pecora da parte dello Staphylococcus aureus sfingomielinasi C (beta-tossina) e un fattore camp22.
Genome mining ha identificato due geni che codificano i fattori CAMP in ogni ceppo Finegoldia sequenziato dal genoma (CAMP1 e CAMP2; Figura S4); ogni omologo ospita il dominio della famiglia dei fattori CAMP (Pfam07373). È interessante notare che un omologo (CAMP2) per ceppo ospita un ulteriore dominio batterico Ig-like (Pfam02368). Oltre a CAMP1 e CAMP2, quattro ” F. i ceppi “nericia” contengono due fattori CAMP aggiuntivi che differiscono sostanzialmente dagli altri omologhi (CAMP3 e CAMP4; Figura S4). Pertanto, quattro ceppi su sette di “F. nericia” ospitano quattro geni del fattore CAMPO.
Successivamente, volevamo sapere se i ceppi di Finegoldia producono una reazione CAMP positiva. Un test della piastra di CAMP agar mostra che alcuni ceppi sono fortemente positivi alla reazione CAMP, mentre per altri ceppi solo poca coemolisi può essere rilevata nelle condizioni di coltivazione applicate a due e cinque giorni di incubazione (Fig. 5, Figura S5). Tutti e tre testati F. ceppi magna erano fortemente CAMPO reazione positiva. Tra i ceppi “F. nericia” abbiamo rilevato una certa eterogeneità; due ceppi erano fortemente positivi alla reazione CAMP, quattro ceppi erano solo leggermente coemolitici e un ceppo, CCUG54800, era chiaramente negativo alla reazione CAMP. L’analisi genomica ha rilevato che nel ceppo CCUG54800 il gene codificante CAMP1 conteneva un inserimento di un locus codificante la trasposasi nell’estremità 5’del gene, che interrompe il gene (Figura S6).

Reazione CAMP di ceppi Finegoldia. Sono state utilizzate piastre di agar TSA con sangue di pecora al 5%. Staphylococcus aureus è inoculato nella striscia centrale. Un test CAMP positivo è indicato dalla lisi completa degli eritrociti all’interfaccia del Finegoldia sp. e le strisce di S. aureus. Tutti e tre i ceppi di F. magna (in rosso) e due dei sette ceppi di “F. nericia” (in blu) hanno mostrato una forte reazione positiva di CAMP dopo due giorni di incubazione anaerobica.
I nostri risultati indicano che i fattori campo di Finegoldia sp. può essere funzionale e che ci sono differenze specifiche del ceppo nell’attività del fattore CAMPO.
Due sistemi di test biochimici non possono distinguere univocamente F. magna da”F. nericia”
Reazioni biochimiche di GPAC inclusi ceppi di Finegoldia sp. sono stati descritti; solo poca variazione biochimica tra Finegoldia sp. ceppi precedentemente segnalati2, 7. Abbiamo testato due sistemi di test biochimici disponibili in commercio che vengono utilizzati per l’identificazione di anaerobi. Il kit “API® 20A” contiene 21 test, tra cui 16 test per l’utilizzo dei carboidrati. Come riportato in precedenza7, nessuno dei carboidrati potrebbe essere metabolizzato da qualsiasi Finegoldia sp. ceppo (dati non mostrati). Anche le altre reazioni (indolo, ureasi, catalasi, idrolisi dell’esculina e liquefazione della gelatina) non hanno rivelato alcuna differenza tra i ceppi di F. magna e “F. nericia” (dati non mostrati). Il kit “RapIDTM ANA II” contiene 18 test per le attività enzimatiche. Mentre gli enzimi saccarolitici sono carenti, tutti i Finegoldia sp. i ceppi producono enzimi proteolitici (Tabella S5). La reazione che rilevava l’attività della fosfatasi era l’unica reazione che differiva tra i ceppi, con tutti i ceppi di F. magna apparentemente negativi e la maggior parte dei ceppi di “F. nericia” positivi. Tuttavia, l’interpretazione di questo risultato del test è limitata poiché il cambiamento di colore era sul bordo del significato. In conclusione, due sistemi di test biochimici commerciali esistenti, API® 20 A e RapIDTM ANA II, non possono distinguere inequivocabilmente F. magna da”F. nericia”. Le proprietà biochimiche di Finegoldia sp. devono essere ulteriormente esplorati con ulteriori test biochimici.
La microscopia rivela differenze superficiali tra F. magna e”F. nericia”
Nel tentativo di visualizzare la morfologia e la struttura superficiale di F. magna e “F. nericia ” abbiamo usato la microscopia elettronica a scansione (SEM) e la microscopia a forza atomica (AFM) su ceppi di F. magna (07T609, 08T492 e 09T408) e “F. nericia” (09T494, 12T272 e 12T306). SEM ha rilevato differenze superficiali: le cellule di F. magna erano più aderenti o aggregative, apparentemente producendo una matrice polimerica extracellulare (Fig. 6). Ciò conferma il lavoro precedente che ha visualizzato una matrice di esopolisaccaridi nei biofilm di F. magna (ceppo FmBs12) mediante microscopia elettronica a scansione e microscopio a scansione laser confocale23. Nella testata ” F. nericia ” ceppi non siamo riusciti a identificare una tale matrice; le cellule sembrano meno aggregative.
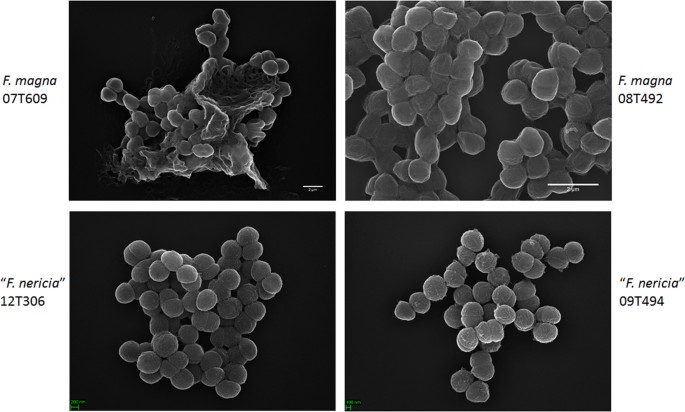
Confronto al microscopio elettronico a scansione di F. magna e “F. nericia”. I pannelli superiore e inferiore mostrano immagini rappresentative di F. magna 07T609 e 08T492 e “F. nericia” 09T494 e 12T306, rispettivamente. Le cellule dei ceppi di F. magna sono più aderenti / aggregative e sembrano produrre una matrice polimerica, in particolare il ceppo 07T609; al contrario, le cellule di “F. nericia” sono meno aggregative.
AFM ha confermato queste osservazioni. Inoltre, abbiamo osservato differenze riguardanti le appendici cellulari. Le cellule di F. magna producevano appendici filamentose più lunghe che sporgevano dall’intera superficie cellulare; queste strutture potevano rappresentare pili o fimbrie (Fig. 7). Al contrario, le cellule dei ceppi di “F. nericia” avevano una superficie più liscia senza lunghe appendici sporgenti. Non si può escludere, tuttavia, che queste osservazioni AFM dipendano dalle condizioni di coltivazione batterica e dalle fasi di lavorazione del campione.

Confronto microscopico della forza atomica di F. magna e “F. nericia”. I pannelli sinistro e destro rappresentano immagini di F. magna 07T609 e” F. nericia ” 12T306, rispettivamente. (A e C) Immagini ad alta risoluzione con metodo soft-touch avanzato assistito con barre di scala Z fino a 0,3 µm e barre XY che indicano 500 nm. (B e D) Immagini sovrapposte di immagini ad alto contrasto di (A e C), combinate con rimozione polinomiale orizzontale e verticale indipendente di (A e C), risolte all’11 ° grado. Gli esperimenti sono stati eseguiti su tre campioni di ciascun ceppo.