- Genomas de estirpes de Finegoldia
- Divisão de Finegoldia cepas em dois distintos subtipos com base no genoma de comparação
- Cepa-específicos heterogeneidade de Finegoldia sp
- factores de interacção do hospedeiro e factores de virulência (putativos)
- os genomas de Finegoldia codificam factores de campo que são funcionais
- dois sistemas de testes bioquímicos não podem diferenciar inequivocamente F. magna de”F. nericia”
- Microscopia revela diferenças de superfície entre F. magna e “F. nericia”
Genomas de estirpes de Finegoldia
No total, 17 de genomas de Finegoldia sp. foram analisados e comparados. Dez estirpes foram recentemente sequenciadas pelo nosso grupo 15; foram isoladas de doentes com infecções ortopédicas associadas ao implante articular na Suécia. Sete genomas foram previamente sequenciados por outros, incluindo o genoma fechado da estirpe do tipo ATCC 2932814. As características genómicas de todas as estirpes sequenciadas de Finegoldia estão resumidas na Tabela 1. A anotação usando Prokka detectou entre 1570 e 1906 sequências de codificação (CDS) por genoma, com uma média de 1760 CDS por estirpe.
Divisão de Finegoldia cepas em dois distintos subtipos com base no genoma de comparação
Nós já notou uma grande heterogeneidade entre os genomas de Finegoldia isolates15. Um alinhamento núcleo-genoma de todos os 17 genomas foi feito e polimorfismos nucleotídicos únicos (SNPs) no genoma central foram chamados. No total, 126.647 núcleos-genoma SNPs foram identificados e usados para a reconstrução filogenômica, revelando que os 17 genomas podem ser agrupados em dois clados distintos (Fig. 1). Uma análise adicional que determina a identificação média do nucleótido (ANI) do genoma do núcleo mostrou que os dois clados exibem um ANI de 90,7%, com uma variação do ANI específico da estirpe de 90,2% para 91,2%. Um ANI tão baixo indica que os dois clados representam especialidades individuais 16. Um clado contém estirpes mais previamente sequenciadas e estudadas, incluindo ATCC29328 e ALB8; pode ser considerado como o clado clássico de F. magna, com um ANI intraclade de 96,6% (variação 95,1–97,4%). O outro clado com um ANI intraclade de 94,8% (93,4–96.6%) contém outra estirpe ATCC (ATCC 53516) e, principalmente, estirpes isoladas de doentes do Hospital Universitário Örebro, Sweden15. Daqui em diante, estirpes deste clado são tentativamente chamadas de “Finegoldia nericia” (latim para Närke, a região Sueca, onde a maioria das estirpes desta espécie foram isoladas).

Phylogenomic tree based on core-genome SNPs of all 17 sequenced genomes of Finegoldia strains. O programa Parsnp foi usado para alinhar o genoma Central e chamar SNPs. O genoma do núcleo é representado por 43% do genoma de referência (ATCC29328); um total de 126.647 SNPs confiáveis do núcleo foram usados para reconstruir uma filogenia do genoma inteiro. Genomas podem ser atribuídos a dois clados principais; um clado representa estirpes de F. magna e o outro clado consiste de estirpes de uma nova espécie, aqui provisoriamente chamado de “F. nericia”.
Uma comparação das sequências de rRNA 16S de todos os isolados revelou um alto grau de semelhança geral (>99%), mas detectou uma separação de F. magna isolados de “F. nericia” isolados, devido à 5 a 9 “F. SNPs específicos da nérica (figura S1). Também revelou uma maior diversificação entre os isolados de “F. nericia” em comparação com F. magna. Em seguida, analisamos sequências adicionais 16S rRNA pertencentes à Finegoldia sp. armazenado em GenBank (figura S2). This revealed that most stored sequences cluster with “F. nericia”, in particular a group of Finegoldia sp. estirpes isoladas de biofilms17. Mais uma vez, uma maior diversificação entre isolados de “F. nericia” é detectada. Em seguida, analisamos os dados microbiológicos humanos armazenados no banco de dados IMG (Integrated Microbial Genomes and microbiome samples) para a presença de sequências de F. magna e “F. nericia”. 3287 e 7068 seqüências de codificação de F. magna ATCC29328 e “F. nericia” ATCC 53516, respectivamente, poderiam ser identificados em 875 montado microbiano humano conjuntos de dados presentes na IMG (dados não apresentados); maior ocorrências de Finegoldia foram detectados no sulco retroauricular amostras. Em conjunto, estas análises sugerem que entre os isolados de Finegoldia associados ao ser humano “F. nericia” parece ser mais prevalente do que F. magna.
Cepa-específicos heterogeneidade de Finegoldia sp
Para lançar mais luz sobre a estrutura populacional do Finegoldia gênero, análises comparativas, de 12 de genomas (todos os dez recém-seqüenciado cepas e as duas cepas ATCC) foram realizadas, incluindo quatro cepas de F. magna clado e oito cepas do “F. nericia” clado. Uma comparação visual dos 12 genomas destacou a diferença de identidade nucleotídica entre os dois clados (Fig. 2). Curiosamente, o plasmídeo encontrado no ATCC 29328 parece ser específico para esta estirpe, uma vez que nenhuma outra estirpe sequenciada contém uma sequência semelhante (Fig. 2A).

o Genoma de comparação, de 12 de estirpes pertencentes ao gênero Finegoldia. Os dois anéis mais íntimos representam o conteúdo de G + C (preto) e o GC-skew (violeta/verde). (A) the reference strain is ATCC29328 (F. magna); note the high nucleotide identity (visualized by the dark red color) of the three innermost genomes (belonging all to F. magna), e o menor identificar oito estirpes pertencentes à “F. nericia” (vermelho pálido cor dos oito anéis exteriores); de dentro para fora: cepas 07T609, 08T492, 09T408, 09T494, 12T272, 12T273, 12T306, CCUG 54800, T151023, T160124 e ATCC 53516. A grande região específica do ATCC29328 na região superior esquerda representa o pFMC plasmídeo. B) a estirpe de referência é 09T494 (“F. Americana”). Os sete genomas “F. nericia” ( anéis internos) têm um nucleótido maior identificado (representado pela cor vermelha mais escura), em comparação com os quatro genomas “F. magna” (anéis exteriores); de dentro para fora: cepas 12T272, ATCC 53516, 12T306, 12T273, CCUG 54800, T151023, T160124, ATCC29328, 07T609, 08T492 e 09T408. Não há sinais de regiões genômicas específicas de “F. nericia”, mas note Os cinco grandes clusters específicos da estirpe.
não foram detectadas diferenças significativas no tamanho do genoma e no número de CDS entre os dois clades. Além disso, nenhuma região genômica específica do clado (ou seja, F. magna ou “F. nericia”-específica) foi detectada, como julgado a partir da análise BRIG (Fig. 2) e também confirmado por uma abordagem bidirecional de explosão de todos os CDS utilizando o ProteinOrtho (quadro S1). Esta abordagem detectou 1202 ortólogos compartilhados por todos os 12 genomas, ou seja, em média 68% dos CDS de cada estirpe fazem parte do proteoma Central. Apenas poucos genes específicos do clado são encontrados. Em vez disso, muitas regiões partilhadas por apenas algumas estirpes podem ser encontradas, por exemplo, para a estirpe 09T494 (ver também Fig. 2B). Além disso, estão presentes regiões específicas da estirpe que codificam no total os CDS específicos da estirpe 1016, com uma grande gama entre as estirpes, por exemplo, 187 e 22 CDS específicos da estirpe são detectados nas estirpes ATCC29328 e T151023, respectivamente (quadro S1). Entre as funções específicas da estirpe estão várias funções de transporte (ferro, bombas de efluxo, transportadores oligopéptidos ABC), proteínas de superfície, sistemas de modificação de restrições, funções de transferência conjugativa, síntese de policetida, bacteriocinas, funções relacionadas com fagos, determinantes da resistência aos antibióticos e sistemas CRISPR/cas.
curiosamente, foram detectadas diferenças nos sistemas CRISPR/cas entre estirpes de F. magna e “F. nericia” (quadro S2). Sete em nove F. as estirpes magna continham um a quatro conjuntos CRISPR por estirpe, contendo, em média, 12 espaçadores (que variam entre 3 e 38 espaçadores por estirpe). A repetição “GTTTGAGAATGATGTAATTTCATAGGTATTAAAC” foi específica para estirpes de F. magna. Em contraste, as estirpes “F. nericia” transportavam apenas uma matriz CRISPR por estirpe que continha em média 53 espaçadores (intervalo 14 a 87 por estirpe). Cada estirpe continha um conjunto individual de espaçadores na(s) SUA (s) matriz (S) CRISPR, heterogeneidade específica da estirpe subjacente, e indicando que cada estirpe tem uma história evolutiva única.
a fim de avaliar se essas funções específicas da estirpe foram adquiridas horizontalmente olhamos para os sinais de sua mobilidade/aquisição usando o programa IslandViewer (figura S3). Este programa previu 141 (estirpe 09T408) a 408 (estirpe 12T273) genes localizados em Ilhas genómicas susceptíveis de serem adquiridas horizontalmente, incluindo muitos dos genes específicos da estirpe acima mencionados envolvidos na especialização, fitness, Sobrevivência e competição interespécie (tabela S3).
factores de interacção do hospedeiro e factores de virulência (putativos)
vários factores de interacção do hospedeiro foram descritos para F. magna 4,5. Foram identificados pili dependentes da Sortase que são importantes para a adesão e colonização13. Um locus genômico que codifica os genes para a subunidade pilus Fmp1, a subunidade putativa Fmp2 e três sortases é conservada entre todos os genomas sequenciados(Fig. 3A). Existem variações específicas da estirpe para cima e para baixo deste locus, i.e. algumas estirpes possuem genes adicionais a montante de fmp1 e a jusante do locus da sortase, incluindo um gene que codifica uma proteína da família hemolisina III. Curiosamente, existem diferenças específicas em relação ao fmp1. Quase cada estirpe carrega uma variante individual e uma análise filogenética de Fmp1 em nível proteico não mostrou nenhuma correlação com a análise filogenômica do genoma do núcleo (Fig. 3B). Em contraste, o Fmp2 é altamente conservado entre todas as estirpes. Outra proteína, codificada a montante do fmp1, abriga domínios de ligação do colagénio e do fibrinogénio; o domínio de ligação ao colagénio (COG4932) também está presente no Fmp2. Propomos que esta proteína também pertença ao pilus locus e nomeie o gene correspondente fmp3.

loci genómico dependente da Cortase nos genomas das estirpes de Finegoldia. (A) Genomic organization of the loci in F. magna (red) and “F. nericia” (light blue) strains. Os três genes de sortases são representados em verde claro (rotulado “2”); o gene para a subunidade pilus principal Fmp1 é mostrado em vermelho (rotulado “1”) e fmp2 é representado em laranja (rotulado”3″). Upstream of fmp1 and downstream of the sortase genes in some strains additional genes (rotulado “8”, hemolysin III-family protein and “10”, hypothetical protein) are present. Uma proteína de ligação ao colagénio putativo, Fmp3 (rotulado “4”), é codificada a montante do fmp1. B) árvore filogenética de fmp1 de estirpes de Finegoldia. A maioria das estirpes possuem uma variante fmp1 individual. A história evolutiva foi inferida usando o método de máxima probabilidade, feito em MEGA7.
a proteína L é provavelmente o factor mais bem estudado de F. magna 8. É um superantigeno que atinge as células B e as imunoglobulinas. Foi detectado em cerca de 10% dos isolados Finegoldia testados até agora, mas varia em tamanho, e número de fragmentos de ligação antigênica (Fab)-domínios de ligação (domínio B1, pfam02246)4,8,18. Uma pesquisa blástica com a proteína l estudada da estirpe 312 (ID locus: M86697, 719 aa) revela algumas proteínas altamente similares (identidade proteica acima de 50%) com diferentes tamanhos (603 a 1417 aa) em algumas estirpes, ou seja, cepas CCUG54800, T161024, ATCC53516, 12T273 e 12T306 (Fig. 4A). Todas estas são estirpes de “F. nericia”, o que indica que o superantigeno das células B é mais prevalente nesta espécie. O fator L da proteína excepcionalmente grande (1417 aa) na estirpe 12T306 possui, além de dois domínios B1, também dois módulos GA que são conhecidos das proteínas de ligação à albumina peptostreptocócica (ver abaixo). Muitas proteínas adicionais com baixa semelhança com a proteína L, mas sem conter quaisquer domínios B1 aparentes, são codificadas nas estirpes de Finegoldia sequenciadas pelo genoma (tabela S4A).; mais uma vez, existem variações específicas da estirpe e algumas estirpes, ou seja, 09T408, 09T494 e T151023, não possuem tais homólogos.

loci genómico que codifica proteínas L e homólogos FAF em estirpes de Finegoldia. A) as pesquisas blásticas com a proteína l caracterizada da estirpe 312 (ID locus: M86697) foram realizadas em estirpes de Finegoldia sequenciadas pelo genoma, e os homólogos (a vermelho, rotulados como “1”) com elevada semelhança (>50% de identidade proteica) são apresentados (ver também Quadro S4A). Apenas estirpes de “F. nericia”, mas não de ” F. as estirpes de magna contêm tais homólogos com diferentes comprimentos. B) as pesquisas por explosão com a proteína FAF da estirpe ALB8 estudada encontraram homólogos de comprimento total (a vermelho, rotulados “1”) em três estirpes de F. magna (a vermelho) e quatro de F. nericia (a Azul claro). O gene FAF é moldado nas duas estirpes” F. nericia ” T151023 e 12T272.
descobriu-se que Finegoldia magna se ligava aos albumin9 humanos. A proteína de ligação à albumina peptostreptocócica (PAB) foi estudada na estirpe ALB8; a sua actividade está ligada a um rate9 de crescimento acelerado. Proteína PAB da estirpe ALB8 (ID locus): X77864) contém módulos GA, o domínio de ligação à albumina; a estrutura do módulo GA foi resolvida 19. Uma busca por explosão revelou que um homolog PAB próximo não pode ser encontrado em outros genomas Finegoldia sequenciados (tabela S4B). Em vez disso, existem vários homólogos que compartilham até 50% da identidade proteica; similaridades de homólogos em estirpes de F. magna são, em média, mais elevadas em comparação com homólogos em estirpes de “F. nericia”. No que diz respeito às proteínas contendo módulos GA, no total 16 proteínas são codificadas nos 12 genomas de Finegoldia, indicando alguma redundância das proteínas ligantes à albumina (tabela S4C). A maioria das estirpes codificam duas proteínas com módulos GA, excepto da estirpe 09T408 (F. magna) e T151023 (“F. nericia”).
a protease da subtilisina ligada à parede celular demonstrou degradar várias proteínas humanas, tais como o colagénio IV,o fibrinogénio e os peptídeos antimicrobianos,tais como LL-37 e MIG/CXCL910, 20, 21. As pesquisas blásticas determinaram que a maioria das estirpes de Finegoldia transportam um homolog SufA altamente semelhante (identidade proteica > 75%) ao SufA estudado da estirpe ALB8 (ID locus: DQ679960). As excepções são as estirpes 07T609, 09T408 e CCUG54800. Um segundo homolog SufA está presente na maioria das estirpes; tem uma identidade proteica média de 26% (tabela S4D).Outro fator importante de interação hospedeira é FAF (fator de adesão de F. magna). Cerca de 90% das estirpes de F. magna produzem este factor, que é responsável pela aglomeração de bactérias e mediata a ligação à membrana basilar, ligando-se ao BM-4011. Descobrimos que sete (três F. magna e quatro estirpes “F. nericia”) das 12 estirpes codificam um homolog FAF semelhante ao estudado na estirpe ALB8 (Fig. 4B). Existem diferenças específicas da estirpe e as mutações dos mecanismos de transição são observadas nas estirpes “F. nericia” T151023 e 12T272. Não se encontra homolog nas estirpes 09T408, 09T494 e 12T306 (quadro S4E).
os genomas de Finegoldia codificam factores de campo que são funcionais
procurámos no genoma outros factores putativos que interagem hospedeiros e encontrámos genes que codificam factores de Christie-Atkins-Munch-Petersen (CAMP). Os factores do campo podem actuar como co-hemolisinas e explicar a chamada reacção do campo, a lise sinérgica dos eritrócitos de ovinos por Staphylococcus aureus sphingomielinase C (beta-toxina) e um campo factor22.
Genoma de mineração identificados dois genes de codificação de ACAMPAMENTO fatores em cada genoma seqüenciado Finegoldia tensão (CAMP1 e CAMP2; Figura S4); cada homóloga abriga o ACAMPAMENTO fator de domínio da família (Pfam07373). Curiosamente, um homolog (CAMP2) por estirpe contém um domínio IG bacteriano adicional (Pfam02368). Além do CAMP1 e do CAMP2, quatro ” F. as estirpes “nericia” contêm dois factores adicionais do campo que diferem substancialmente dos outros homologs (CAMP3 e CAMP4; figura S4). Assim, quatro de sete estirpes “F. nericia” abrigam quatro genes do fator de campo.A seguir, queríamos saber se as estirpes de Finegoldia produzem uma reacção positiva no campo. Um ensaio em placa de ágar do campo mostra que algumas estirpes são fortemente positivas à reação do campo, enquanto para outras estirpes apenas pode ser detectada pouca Co-hemólise nas condições de cultivo aplicadas em dois e cinco dias de incubação(Fig. 5, Figure S5). Todos os três testaram F. as estirpes de magna eram fortemente positivas. Entre as estirpes de “F. nericia” detectámos alguma heterogeneidade; duas estirpes eram fortemente positivas para a reacção do campo, quatro estirpes eram apenas ligeiramente Co-hemolíticas, e uma estirpe, CCUG54800, era claramente negativa para a reacção do campo. A análise genômica descobriu que na estirpe CCUG54800 o gene de codificação CAMP1 continha uma inserção de um locus de codificação de transposase na extremidade 5’do gene, que perturba o gene (figura S6).

reacção de campo das estirpes de Finegoldia. Foram utilizadas placas de ágar TSA com 5% de sangue de ovelha. Staphylococcus aureus é inoculado na faixa do meio. Um teste de campo positivo é indicado por lise eritrocitária completa na interface da Finegoldia sp. e as riscas de S. aureus. Todas as três estirpes de F. magna (a vermelho) e duas de sete estirpes de F. nericia (a azul) mostraram uma forte reação positiva do campo após dois dias de incubação anaeróbica.
os nossos resultados indicam que os factores de campo da Finegoldia sp. pode ser funcional e que existem diferenças específicas da estirpe na actividade do factor do campo.
dois sistemas de testes bioquímicos não podem diferenciar inequivocamente F. magna de”F. nericia”
reacções bioquímicas da GPAC incluindo estirpes de Finegoldia sp. foram descritas apenas pequenas variações bioquímicas entre a Finegoldia sp. as estirpes foram previamente notificadas2,7. Testamos dois sistemas de testes bioquímicos disponíveis comercialmente que são usados para a identificação de anaeróbios. O kit “API® 20A” contém 21 testes, entre os quais 16 testes de utilização de carboidratos. Como relatado anteriormente 7, nenhum dos carboidratos pode ser metabolizado por qualquer Finegoldia sp. estirpe (dados não apresentados). As outras reacções (indole, urease, catalase, hidrólise esculina e liquefação de gelatina) também não revelaram qualquer diferença entre estirpes de F. magna e F. nericia (dados não apresentados). O kit “RapIDTM ANA II” contém 18 testes para actividades enzimáticas. Enquanto faltam enzimas sacarolíticas, todas as Finegoldia sp. as estirpes produzem enzimas proteolíticas (quadro S5). A reação que detectou a atividade da fosfatase foi a única reação que diferiu entre as estirpes, com todas as estirpes de F. magna aparentemente negativas e a maioria das estirpes de “F. nericia” positivas. No entanto, a interpretação deste resultado de teste é limitada, uma vez que a mudança de cor estava no limite de significância. Em conclusão, dois sistemas de teste bioquímico comercial existentes, API® 20 A E RapIDTM ANA II, não podem diferenciar inequivocamente F. magna de “F. nericia”. The biochemical properties of Finegoldia sp. precisa ser mais explorado com testes bioquímicos adicionais.
Microscopia revela diferenças de superfície entre F. magna e “F. nericia”
numa tentativa de visualizar a morfologia e a estrutura superficial de F. magna e ” F. nericia ” we used scanning electron microscopy (SEM) and atomic force microscopy (AFM) on strains of F. magna (07T609, 08T492, and 09T408) and “F. nericia” (09T494, 12T272 and 12T306). SEM detectou diferenças de superfície: as células de F. magna eram mais aderentes ou agregantes, aparentemente produzindo uma matriz polimérica extracelular (Fig. 6). Isto confirma trabalhos anteriores que visualizaram uma matriz de exopolissacarídeos em biofilmes de F. magna (estirpe FmBs12) por varredura de microscopia eletrônica, bem como microscopia de varredura de laser confocal23. In the tested “F. nericia ” estirpes que não conseguimos identificar tal matriz; as células parecem menos agregadas.
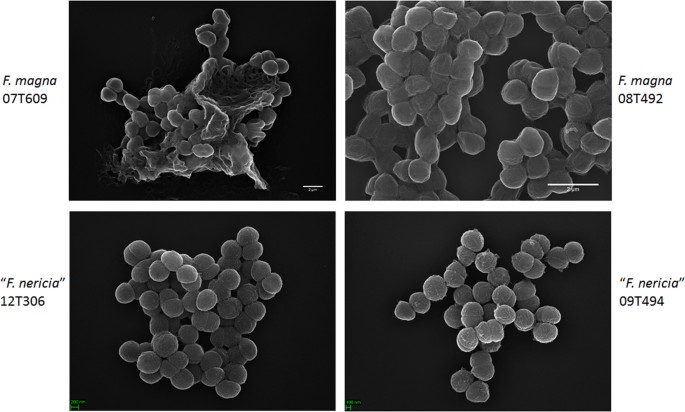
Scanning electron microscópica comparação de F. magna e “F. nericia”. Os painéis superior e inferior mostram imagens representativas de F. magna 07T609 e 08T492 e” F. Americana ” 09T494 e 12T306, respectivamente. As células de estirpes de F. magna são mais aderentes / agregantes e parecem produzir uma matriz polimérica, em particular a estirpe 07T609; em contraste, as células de “F. nericia” são menos agregadas.
a AFM confirmou estas observações. Além disso, observamos diferenças em relação aos apêndices celulares. Células de F. magna produziram apêndices filamentosos mais longos que se projetavam a partir de toda a superfície celular; estas estruturas poderiam representar pili ou fimbriae(Fig. 7). Em contraste, as células de estirpes de” F. nericia ” tinham uma superfície mais suave, sem longos apêndices salientes. Não pode ser descartado, no entanto, que estas observações AFM dependem das condições de cultivo bacteriano e das etapas de processamento da amostra.

de força Atômica microscópica comparação de F. magna e “F. nericia”. Os painéis esquerdo e direito representam imagens de F. magna 07T609 e” F. nericia ” 12T306, respectivamente. (A E C) imagens de alta resolução com método soft-touch avançado assistido com barras de escala Z até 0,3 µm e barras XY indicando 500 nm. (B E D) sobrepor imagens de alto contraste de (A E C), combinadas com remoção polinomial horizontal e vertical independente de (A E C), resolvidas até o 11º grau. Foram realizadas experiências em três amostras de cada estirpe.